
Rhodolith (Coralline Red Algae) beds are ubiquitous sea-floor ecosystems distributed all over the world. Equivalent dense to loose concentrations of Rhodoliths, as major components of carbonate, are also common in the rock record in tropical and cold-temperate settings. They can be the result of the accumulation of Rhodoliths where they originally grew (autochthonous concentrations), or nearby (parautochthonous Rhodolith beds), or can be accumulated in a different site due to long-distance transport (allochthonous concentrations). Reworked deposits can accumulate either offshore or onshore of the original place of Rhodolith development. The oldest records of calcified Coralline Algae in the Early Cretaceous are fragments dispersed in the sediment or thin crusts attached to Corals. Coralline Algae became important carbonate components during the latest EarlyCretaceous, and Rhodoliths were increasingly common during the Late Cretaceous. There are no quantitative studies on the volumetric contribution of Rhodolith beds to the global carbonate production through time. Qualitative data have only been compiled for Oligocene–Neogene Coralline Algal-dominated deposits. Although most of this information is based on the Mediterranean record, the data suggest an important increase in Rhodolith bed occurrences during the Oligocene and early Miocene. Interestingly, this substantial development coincidewith the highest Coralline diversity in the Chattian–Burdigalian (Late Oligocene-Early Miocene, 28.1-18.97 million years ago) interval. Global temperature, palaeoceanographic circulation patterns, and major palaeogeographic changes in the Tethys were important factors controlling the diversification of Coralline Algae. Before this diversity peak, during the Eocene, Rhodoliths were widespread worldwide, but information on their relevance in marine ecosystems is scarce. The Eocene was a transient period of time between greenhouse and icehouse conditions. Global temperature rose in the early Eocene up to the highest values ever reached during the Cainozoic, during the Early Eocene Climatic Optimum. Then, temperature progressively decreased during the rest of the Eocene, the Long-term Eocene Cooling, up to a drastic cooling at the Eocene–Oligocene transition due to the onset of the Southern Hemisphere glaciation.
In a paper published in the journal Diversity on 23 March 2020, Julio Aguirre and Juan Braga of the Departamento de Estratigrafía y Paleontología at the Universidad de Granada, Victoriano Pujalte and Xabier Orue-Etxebarria at the Departamento de Estratigrafía y Paleontología at the Universidad del País Vasco, Edward Salazar-Ortiz of the Servicio Geológico Colombiano, Daniel Rincón-Martínez of the Ecopetrol Centro de Innovación y Tecnología, Manuel Abad of the Departamento de Biología y Geología, Física y Química Inorgánica at the Universidad Rey Juan Carlos, and Fernando Pérez-Valera of the Departamento de Ciencias de la Tierra y del Medio Ambiente at the Universidad de Alicante, present the results of a study of Middle Eocene Rhodoliths from four areas, two in Southern Spain, one in Hispaniola (Dominican Republic), and one in Colombia, made in order to analyze the palaeonvironmental conditions in which Rhodoliths developed in different palaeolatitudinal contexts. This will also contribute to understanding the long-term role of coralline algae in marine ecosystems during a time interval when substantial climate changes were taking place at global scale.
Parallel to high global temperatures, sea level several tens of meters higher than in the modern oceans prevailed during the early and middle Eocene, promoting the expansion of large carbonate platforms. In this context, oceans were devoid of large coral reef structures, particularly affecting low latitude areas; the so-called Early–Middle Eocene 'reef gap'. In contrast, large carbonate ramps largely dominated by larger benthic Foraminifers and Algae developed.
As far as is known global diversity of Coralline Algae was moderate during the Eocene, predating the great diversification event during the Oligocene–Early Miocene. During the Middle Eocene, Coralline Algal diversification shows a slight decline coinciding with a major macroevolutionary turnover in Algal assemblages, characterised by a substantial increase of members of the Order Hapalidiales and a decrease of members of the Family Sporolithales.
Palaeogeographic reconstructions during the middle Eocene show that Southern Spain was in mid latitudes while Hispaniola and Colombia were in lower latitudes closer to the equator.
Samples with rhodoliths were collected in the different study areas. Except in one of the study areas in Southern Spain (Salinas Menores), sediments were well lithified, precluding the extraction of isolated rhodoliths to make measurements. Therefore, Rhodolith morphology is based on sections in the outcrops. Samples were cut to produce a total of 56 thin sections.
Aguirre et al. studied two sites in the Betic Cordillera: the Salinas Menores Ravine and Sierra del Zacatín









Parallel to high global temperatures, sea level several tens of meters higher than in the modern oceans prevailed during the early and middle Eocene, promoting the expansion of large carbonate platforms. In this context, oceans were devoid of large coral reef structures, particularly affecting low latitude areas; the so-called Early–Middle Eocene 'reef gap'. In contrast, large carbonate ramps largely dominated by larger benthic Foraminifers and Algae developed.
As far as is known global diversity of Coralline Algae was moderate during the Eocene, predating the great diversification event during the Oligocene–Early Miocene. During the Middle Eocene, Coralline Algal diversification shows a slight decline coinciding with a major macroevolutionary turnover in Algal assemblages, characterised by a substantial increase of members of the Order Hapalidiales and a decrease of members of the Family Sporolithales.
Palaeogeographic reconstructions during the middle Eocene show that Southern Spain was in mid latitudes while Hispaniola and Colombia were in lower latitudes closer to the equator.
Samples with rhodoliths were collected in the different study areas. Except in one of the study areas in Southern Spain (Salinas Menores), sediments were well lithified, precluding the extraction of isolated rhodoliths to make measurements. Therefore, Rhodolith morphology is based on sections in the outcrops. Samples were cut to produce a total of 56 thin sections.
Aguirre et al. studied two sites in the Betic Cordillera: the Salinas Menores Ravine and Sierra del Zacatín

(A) Geographic location of the study areas in Southern Spain (red rectangles). (B) Location of the Salinas Menores Ravine section (blue star). (C) Location of the Sierra del Zacatín area (blue star). Red lines indicate the main roads; blue lines and patch mark rivers and water reservoir, respectively; green lines mark tracks. Aguirre et al. (2020).
The Salinas Menores outcrop is close to Dehesas de Guadix and Villanueva de las Torres villages (Northeast Granada province, Southern Spain). The study deposits belong to the Cañada Formation, which includes redeposited materials due to gravitational flows unconformably overlying different Paleogene–Mesozoic rocks. The section consists of grey-greenish marls intercalating turbidite beds of packstone–rudstone. The marls also incorporate large, up to hundred metre olistoliths derived from the Paleogene–Mesozoic basement. The top of the section is a channelized body, up to 3 m thick, of rudstones. Rhodoliths are dispersed in the marls. Planktic Foraminifera assemblages are characterized by Subbotina jacksonensis, Subbotina senni, Catapsydrax unicavus, Acarinina bullbrooki, Acarinina punctocarinata, Acarinina praetopilensis, Morozovelloides bandyi, Morozovelloides crassatus, and Globigerinatheka spp. This assemblage indicates a Lutetian age (Middle Eocene, 47.8-41.2-million-years-old).

Stratigraphic column from Salinas Menores ravine. Aguirre et al. (2020).
The Sierra del Zacatín is a southwest–northeast trending mountain range northeast of Nerpio (Albacete) in Southern Spain. The study section is located at the southwest of the sierra. The Sierra del Zacatín is mostly made up of Paleocene to Eocene sediments. The middle Eocene succession consists of about 40 m thick limestones dominated by larger benthic Foraminifers, followed by fragments of Echinoids, Bryozoans, and Bivalves. In the middle of the section, Corals become common together with Coralline Algae, which occur as Rhodoliths but are mostly intergrown with Corals. The presence of Alveolina boscii, Alveolina stercusmuris, and Alveolina praespira at the base of the limestones, as well as Alveolina fusiformis, Alveolina elongata, and Nummulites aturicus at the top indicates a Lutetian–earliest Bartonian age (Middle Eocene).

Stratigraphic column from Sierra de Zacatín. Aguirre et al. (2020).
The Dominican Republic shares with Haiti the Hispaniola Island, one of the Greater Antilles separating the Caribbean Sea from the Atlantic Ocean. The Bahoruco Peninsula is the southern extreme of the island and, therefore, the farthest area from the convergence zone of the Caribbean and North America plates, which runs east–west between Hispaniola and the Bahamas. The Cainozoic succession in the Bahoruco Peninsula includes a series of Eocene to Pliocene carbonate units overlying Upper Cretaceous volcanic rocks. The lowest carbonate unit, Polo Unit, consists of whitish limestones, 100 to 250 m thick. The lower part of the unit comprises roughly bedded bioclastic wackestones and Rhodolith rudstones. Coralline Algae are the main components in the latter lithofacies with secondary Corals, Molluscs, and benthic Foraminifera. The larger benthic Foraminifer assemblages include Discocyclina, Lepidocyclina, Nephrolepidina, Operculina, Eorupertia?, Rotalia, Sphaerogypsina, and Cushmania. Upwards in the succession, bedding definition increases as the grain size decreases. The finer-grained limestones in the upper part of the Polo Unit change gradually to wackestones and mudstones included in the Neiba Formation. Larger benthic Foraminifers together with planktic Foraminifers (Acarinina, Globigerina, and Globigerapsis) suggest a Middle (Late) Eocene age.

The inset shows a map of Hispaniola with indication of the study area (red rectangle). Location of the Polo section (blue star). Red lines indicate main roads. Aguirre et al. (2020).
The studied rhodoliths were collected at 18°5’13”N–71°17’33”W, 1 km to the northwest of the hamlet Polo.

Stratigraphic column from Polo. Aguirre et al (2020).
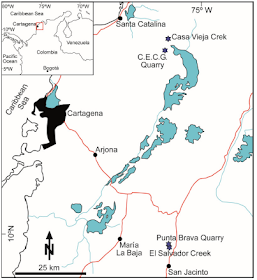
The study Rhodoliths occur in middle Eocene deposits in the San Jacinto Fold Belt, in northwestern Colombia. The San Jacinto Fold Belt is an elongated antiform structure with a northeast–southwest direction. Upper Paleocene-Eocene sedimentary rocks in this belt formed on an intraoceanic Cretaceous Caribbean arc. Two middle Eocene units crop out in the San Jacinto Fold Belt: the Arroyo de Piedra and Chengue formations. Their age assignment is based on planktic Foraminifers and calcareous nannoplankton. The Chengue Formation consists of mudstones, siltstones, and redeposited bioclastic carbonates, up to 300 m in thickness. The Arroyo de Piedra Formation mainly includes bioclastic limestones, sandstones, siltstones, and mudstones.

The inset shows a map of Colombia with indication of the study area (red square). Location of sections (blue stars). Red lines indicate main roads; blue lines and patches mark rivers and lakes, respectively. Aguirre et al. (2020).

Stratigraphic columns from Colombia. Aguirre et al. (2020).
At Salinas Menores Ravine Rhodoliths occur scattered in the planktic foraminifer-rich marls. They are spheroidal to ellipsoidal and range from 2 to 6 cm in largest diameter. Rhodoliths are made up of Coralline Algae intergrown with Bryozoans and encrusting Foraminifers. The Peyssonneliacean Alga Polystrata alba is another common component. The nuclei of Rhodoliths are fragments of Corals, Coralline Algae or wackestone–packstone dominated by Miliolid Foraminifers. On the Rhodolith surface, Algae are predominantly encrusting, with a few warty growth forms. Internally, Rhodoliths are concentric with encrusting Algae followed by warty and fruticose Algal thalli. In terms of Algal assemblages, Rhodoliths are composed of members of the orders Hapalidiales, Sporolithales, and Corallinales. Corallinales is the most diversified group represented by Neogoniolithon spp., Spongites spp., Lithoporella sp., Hydrolithon cf. lemoinei, Subterraniphyllum thomasii, laminar thalli of Karpathia sphaerocellulosa, as well as unidentifiable fragments of geniculate forms showing cell fusions.

(A) Rhodoliths dispersed in planktic Foraminifer-rich marl from Salinas Menores area. (B) Coralline Algal crusts (white veneers) intergrown with Coral from Sierra del Zacatín. (C) Spheroidal Rhodolith made up by the superposition of thin crusts of Coralline algae (Sierra del Zacatín). (D) Autochthonous Rhodolith rudstone from Colombia (CECG Quarry). (E) Redeposited Rhodolith rudstones from Colombia (Punta Brava Quarry). (F) Larger benthic Foraminifer-dominated limestones from Sierra del Zacatín. Aguirre et al. (2020).
At Sierra del Zacatín Coralline Algae mostly occur as thick crusts attached to and intergrowing with Corals. Rhodoliths, although present, occur scattered in the sediment in dispersed and loosely packed Rhodolith beds. Rhodoliths are mainly spheroidal and ellipsoidal, up to 4-5 cm in largest diameter. They are made of Coralline Algae intergrown with encrusting benthic Foraminifers, Bryozoans, Corals, Serpulids, and Vermetids. Other Rhodophytes, such as Parachaetetes asvapatii included in the family Elianellaceae and the Peyssonneliacean Polystrata alba, can be important Rhodolith builders. Algal thalli both on the surface and in the Rhodolith interior are largely encrusting, with very few warty Algal growth forms. A few fruticose thalli are observed in the inner parts of some Rhodoliths.
Coralline Algal assemblages are dominated by Sporolithales, while Hapalidiales and Corallinales are rare.
In the Dominican Republic Rhodoliths occur either densely concentrated in rudstones together with Coralline Algal debris or dispersed in a fine-grained matrix in bioclastic wackestones. They are spheroidal to ellipsoidal and a few centimeters in size. Their internal structure is generally concentric, made up of encrusting to warty thalli although fruticose thalli are also common. Crustose Coralline Algae are intergrown with Peyssonneliaceans, Bryozoans, and encrusting Foraminifers. Rhodoliths are bioeroded by Sponges (Entobia) in varying degrees.
The most abundant components of the Algal assemblages are members of the order Sporolithales followed by Hapalidiales. Members of the order Corallinales are not present in the Dominican material.
At the Colombian outcrops Rhodoliths are the main components in roughly bedded, whitish to light beige rudstones. Coralline Algal fragments, larger and small benthic Foraminifers, and minor fragments of Bivalves and Echinoderms also occur as bioclasts. Rhodoliths also occur in grainstones–packstones to rudstones intercalated in mudstones and siltstones. These lilthofacies appear in decimeter-scale plane-parallel, plane-convex, and channelized beds with rip-up intraclasts of siltstone and mudstone. Red algal fragments are the main components in packstones and in the rudstone matrix (packstone). Geniculate Coralline Algae and larger benthic Foraminifers are also common.
In both types of lithofacies, Rhodoliths are ellipsoidal to spheroidal, and are millimeters to several centimeters in size; they can be fruticose and branching or have concentric to box-work internal structure. The nuclei made up of bioclasts are relatively small compared with the Algal cover (one-fifth to one-sixth in size). Peyssonneliaceans are commonly intergrown with crustose Corallines and occasionally are the only components in some spheroidal nodules. Both kinds of Red Algae are intergrown with encrusting Foraminifers in 30% of nodules.
Coralline Algal assemblages are mainly characterized by members of the orders Hapalidiales and Sporolithales, with anecdotal occurrences of Lithoporella sp.
In the Salinas Menores, Rhodoliths are scattered in the marls, not concentrated in particular beds. The sediment trapped in the inner voids of the Rhodoliths and that surrounding them show different micropalaeontological content. Miliolid Foraminifers are very abundant in the sediment filling up the internal cavities of Rhodoliths. These benthic Foraminifers are typical inhabitants of protected lagoons as well as shallow inner-platform settings. However, the sediment engulfing the Rhodoliths is rich in planktic Foraminifers characteristic of deep environments. The micropalaeontological content suggests that these Rhodoliths are displaced from their original place of growth. The Miliolids trapped within the Rhodoliths when they were growing suggest that the Algal nodules formed originally in a shallow platform. The abundance of members of the order Corallinales is typical of shallow water settings. Rhodoliths were afterwards transported to deeper basinal areas accumulating as allochthonous components. The presence of turbidites, channeled bodies, and olistoliths of varying sizes intercalated in the marls attests that downslope transport was common during the deposition of these materials.
The major biotic components of the middle Eocene carbonates in the Sierra del Zacatín are larger benthic Foraminifers. Coralline Algae are present but as secondary representatives of the fossil assemblages. They occur mainly as crusts, attached to and intergrown with Corals, and as Rhodoliths, which are dispersed in the larger benthic Foraminifer-dominated carbonates. Algal crusts occur on top of Coral colonies of varying sizes, indicating that colonies were preserved in their original growth position. In addition, geopetal structures infilling internal voids of Rhodoliths indicate normal polarity. This is consistent with absence of substantial reworking or lateral displacements of the Algal nodules.
In the lower Polo Unit in the southern Dominican Republic, spheroidal to ellipsoidal Rhodoliths mainly composed of Hapalidiales and Sporolithales indicate relatively deep (several tens of meters) shelf environments. Abundant Peyssonneliacean Red Algae are also characteristic of relatively deep shelf settings. The larger benthic Foraminifers associated to rhodoliths also suggests this kind of shelf environment. Lithofacies changes indicate deepening upwards in the succession.
The Rhodolith rudstones in the San Jacinto Fold Belt in northwestern Colombia probably formed in similar relatively deep-water, mesotrophic shelf environments. Foralgaliths of Hapalidiales and encrusting Foraminifers are also characteristic of calm-water conditions. An increase of Hapalidiales and Peyssonneliaceans in Red Algal assemblages with depth was reported in the Priabonian of Austria, Late Eocene–Late Oligocene in Northeast Italy, and Miocene of Southern Spain. Rhodolith beds dominated by Hapalidiales and Peyssonneliaceans are generally recorded in middle-ramp settings in both modern and ancient depositional systems.
The bed geometries, internal structures, and rip-up clasts clearly indicate that the packstones and rudstones intercalated in mudstones and siltstones in the San Jacinto Fold Belt are sediment gravity flow deposits. The Rhodoliths in this lithofacies were removed from shallower shelf settings and redeposited in deeper marine environments in which the autochthonous sediments were mudstones and siltstones with planktic Foraminifers. Bed geometry and dimensions indicate that these redeposited carbonates accumulated in small channel and lobe systems, downslope of the shelf in which Rhodolith rudstones formed. The Coralline Algal composition does not differ significantly among Rhodolith rudstones from mid-platform and those transported into deeper settings. This suggests that redeposited Rhodoliths originally grew in the same middle platform palaeoenvironments.
The Rhodoliths studied by Aguirre et al. are dominated by Hapalidiales and Sporolithales. Representatives of Corallinales are mostly limited to the pervasive presence of laminar thalli of Lithoporella sp. and calcified segments of geniculates (branching fronds), except in Salinas Menores, where this order is relatively abundant and diverse. Extant species of the order Sporolithales are most diversified in relatively deep tropical waters, although they also occur in shallow waters. Along the evolutionary history of the Coralline Algae, Sporolithales expanded worldwide and reached its highest diversification during the Late Cretaceous, when greenhouse conditions prevailed. Afterwards, the species richness progressively decreased as the planet underwent a general decline in temperature.

Selected Coralline Algal species identified in the different study areas. (A) Sporolithon nummuliticum from the El Salvador Creek section (Colombia). (B) Sporolithon sp. from the Sierra del Zacatín section. (C) Lithothamnion camarasae from the Salinas Menores section. (D. ‘Palaeothamnium’ kossovense from the Polo section. (E) Neogoniolithon sp. 1 from the Salinas Menores section. (F) Lithoporella sp. from the Salinas Menores section. (G) Subterraniphyllum thomasii from the Salinas Menores section. (H) Distichoplax biserialis from the Sierra del Zacatín section. Aguirre et al. (2020).
Hapalidiales, which outnumber other orders in Coralline Algal assemblages in modern cold, high-latitude waters and deeper low-latitude settings, diversified during the Eocene, becoming more abundant than Sporolithales. The Cainozoic decline in temperature started by the end of the Early Eocene and accelerated at the end of this epoch with the onset of glaciation in Antarctica.
In terms of relative abundance, the Coralline Algal assemblages in mid-latitude Southern Spain and in the tropical Dominican Republic and Colombia show varying proportions of Sporolithales and Hapalidiales. The number of species belonging to these two groups varies in the different study areas. In Salinas Menores, Hapalidiales and Corallinales encompass higher species richness than Sporolithales. Complied data from the literature show that Hapalidiales started to diversify in the Ypresian (early Eocene) while Sporolithales slightly declined during the Eocene. Aguirre et al.'s data confirm the increasing replacement of Sporolithales by Hapalidiales during the greenhouse Middle Eocene.
The occurrence of Subterraniphyllum thomasii in the Salinas Menores section is remarkable. This species was particularly abundant during Oligocene times, and some authors have considered it as a biostratigraphic indicator of this epoch. Nonetheless, in the original description of the species, it is indicated that Subterraniphyllum thomasii also rarely occurs in Late Eocene and Aquitanian (Early Miocene) sediments. The presence of Subterraniphyllum thomasii in Salinas Menores extends back its occurrence to the Lutetian (early Middle Eocene).
Distichoplax biserialis is an extinct Coralline Alga characterized by laminar thalli with an isobilateral organization. Distichoplax biserialis is particularly abundant in Palaeocene and Early Eocene carbonates and became gradually extinct during the Eocene. In Aguirre et al.'s study areas, this species is virtually absent except for a few small fragments of thalli found in the Sierra del Zacatín, confirming its rarity in the middle Eocene.
Ecological factors required for the healthy development of Rhodolith beds in recent oceans are well-oxygenated bottom conditions, low sedimentation rates, low content of suspended particles, and moderate water energy. Except where Rhodoliths were transported from shallower settings, Rhodoliths in the rest of the study areas formed in oxygenated conditions, as shown by the prolific abundance of accompanying faunas, such as Sea Urchins, Corals, larger benthic Foraminifers, Bryozoans, and Molluscs. In addition, carbonate sedimentation devoid of terrigenous particles indicates low sediment supply and, consequently, clear waters. Finally, absence of sedimentary structures suggests that turbulence was low to moderate.
Although local palaeoenvironmental conditions were a priori favorable for Rhodolith bed development, Rhodoliths are major constituents in the study tropical Middle Eocene shallow platform deposits, whereas larger benthic Foraminifers with varying proportions of Calcareous Red Algae dominate in carbonate deposits at mid latitude. Similarly, Eocene deposits worldwide are mostly characterised by Rhodoliths and Coralline Algal fragments dispersed in larger benthic Foraminifer-dominated carbonates, and the few examples of Rhodolith beds were found so far in the Early or Late Eocene. The low proportion of densely packed Rhodolith beds during the Eocene, and particularly during the Middle Eocene, coincides with a relative decline in Algal diversity and with a significant decline in reef ecosystems.

Distribution of Rhodolith beds (red circles) and larger benthic Foraminifer-dominated carbonates with variable proportions of Rhodoliths (blue circles) during the Eocene. Diamonds indicate Rhodolith beds (red diamonds) and larger benthic Foraminifer-dominated carbonates with dispersed Rhodoliths (yellow diamonds) but with no chronological precision. Aguirre et al. (2020).
An earlier study performed a detailed facies analysis of Middle Eocene to lower Oligocene ramp carbonates in different localities from Central and Southern Alps. Interestingly, Coralline-Algal dominated facies (marl, Rhodolith, Algal debris, and crustose Algal facies) are frequent in Late Eocene and Early Oligocene deposits, while Middle Eocene carbonate facies were largely dominated by larger benthic Foraminifers with subordinate Algal debris and local Rhodolith concentrations in middle ramp settings.
Likewise, at several seamounts southeast of Japan, Middle–Late Eocene shallow water carbonates mainly dominated by larger benthic Foraminifers have been described. Coralline Algae, forming Rhodoliths or as fragments in rudstones to packstones, occur in lesser abundance. They became dominant afterwards, in Oligocene-to-Pleistocene carbonate deposits in the same western Pacific areas.
Profuse development of larger benthic Foraminifers mostly takes place in oligotrophic conditions, although they can be also important in nutrient-rich tropical sediments in upwelling areas. Regarding Coralline Algae, it is not clear whether nutrient contents do actually promote the development of Rhodolith beds. Present-day Coralline Algae withstand strong annual variations in nutrient conditions, from nearly depleted settings to high levels of nutrients. However, it seems that profuse Rhodolith beds mainly occur in mesotrophic conditions. The largest Rhodolith beds in tropical latitudes occur nowadays on the eastern Brazilian shelf, in areas with relatively reduced development of Coral reefs. Here, Rhodolith beds extend from shallow subtidal settings to the shelf margin and thrive under mesotrophic conditions, with mean seawater temperatures higher than 20°C on the sea floor, and low terrigenous sedimentation, which is generally limited to near-shore areas. Similarly, extensive Rhodolith beds are found in the Amazon River mouth in the northwestern Brazilian platform, associated to the so-called Great Amazon Reef System. This is a complex of carbonate buildups including Scleractinian Corals, encrusting Coralline Algae, Sponges, and Rhodolith beds developed in the marginal areas of siliciclastic influx from the Amazon River under mesotrophic conditions.
In subtropical latitudes in the Gulf of California, Rhodoliths spread throughout the gulf. Large and dense Rhodolith beds extend from shallow subtidal zone to about 40 m depth and occur in a wide spectrum of environmental conditions, with extreme variations of temperature (8–32°C), and in mesotrophic waters in the middle part of the Gulf of California. Fine sediment input and related anoxia seem to be strong limiting factors for Rhodolith development.
In a similar way, the Rhodolith beds in the Mediterranean occur in mesotrophic areas with reduced sedimentation and far from high nutrient influx. In the subtropical Western Pacific, on the shelves around the Ryukyu Islands, Rhodolith beds develop in nutrient poor waters lacking significant upwelling.
The greenhouse conditions prevailing during great part of the Eocene favored the establishment of productive equatorial ocean waters and oligotrophic conditions widespread in middle and high latitudes. Palaeoceanographic models as well as type of sediments show that productive upwelling zones were located in low latitudes, particularly in the Pacific, during the Eocene. There is evidence of low nutrient conditions at midlatitude in the southeastern Atlantic Ocean during the Middle Eocene Climatic Optimum. In Aguirre et al.'s study cases, the Middle Eocene deposits of the Chengue Formation in Colombia were formed in mesotrophic waters according to their micropalaeontological content. Aguirre et al. hypothesise that latitudinal gradient in oceanic productivity might account for the formation of Rhodolith beds and Rhodolith rudstone lithofacies in tropical areas, whereas larger benthic Foraminifer-enriched lithofacies prevailed in mid latitudes.
A precise reconstruction of environmental variables in middle Eocene carbonate records is difficult, which is generally true for Palaeogene larger benthic Foraminifer- and Coralline Algal-dominated sediments. The impact of high temperatures due to high levels of atmospheric carbon dioxide during the Eocene, and particularly during the hyperthermal events, on Rhodolith bed development needs to be assessed. In this regard, sustained anomalously high summer temperatures led to high mortality rates of Coralline Algae in rhodolith communities along the western coast of Australia. In addition, and taking into consideration the discussion made above, the prevailing oligotrophic conditions at global scale accounting for the general prevalence of larger benthic Foraminifers during the Middle Eocene and the relative decline of Rhodolith beds worldwide requires further analyses. More calibration studies, essentially geochemical, for reconstructing water temperature and palaeoproductivity, and knowledge of the depth habitats of benthic and planktic organisms are needed to define the multifactor settings that drove the carbonate grain associations found in low- and mid-latitude regions during the Middle Eocene.
In two tropical settings, the Dominican Republic and Colombia, Middle Eocene Coralline Algae occur as dense concentrations of Rhodoliths in Rhodolith rudstone lithofacies. Rhodoliths are ellipsoidal to spheroidal in shape and are composed by encrusting to warty Coralline Algal thalli in association with benthic Foraminifers, Bryozoans, Corals, and other calcareous Red Algae such as Parachaetetes asvapatii and Polystrata alba.
In mid-latitude areas in Southern Spain, Coralline Algae occur in two different contexts. In Salinas Menores ravine, Rhodoliths are spheroidal to ellipsoidal and occur dispersed in planktic Foraminifer-rich marls. In the Sierra del Zacatín, larger benthic Foraminifers dominate the Middle Eocene carbonate deposits and Rhodoliths are scarce. Coralline Algae mostly occur attached to and intergrown with Corals. Rhodoliths consist of encrusting and warty (occasionally fruticose) Algal thalli intergrown with Bryozoans, Corals, and benthic Foraminifers.
In all the study areas, Coralline Algal assemblages are dominated by Hapalidiales and Sporolithales. The order Corallinales is scarcely represented, except in Salinas Menores, where its members are relatively abundant and diverse. Within this group, Subterraniphyllum thomasii is found in the Salinas Menores section. The oldest previously known record of this species is from the Late Eocene; therefore, Aguirre et al.'s finding extends the occurrence of Subterraniphyllum thomasii back to the Middle Eocene.
Rhodoliths in the two tropical areas developed in relatively deep platform settings (tens of meters of water depth) as shown by the larger benthic Foraminifers and Coralline Algal assemblages. In the Salinas Menores section, Miliolids in the internal voids indicate that Rhodoliths grew in a shallow-water inner platform or lagoon and were re-deposited in deep outer-shelf settings.
During the greenhouse conditions in the Early–Middle Eocene, shallow-water carbonate platforms from the tropics to intermediate latitudes were depauperate in Rhodolith beds. The key palaeoenvironmental factors accounting for this decline remain elusive up to now. Extremely high global temperatures due to high atmospheric carbon dioxide concentrations could negatively affect Coralline Algae. In addition, distribution of oceanic productivity might account for the main carbonate producers in marine shelves: mesotrophic conditions associated with upwelling areas in tropical regions could have favored the development of Rhodolith beds, such as those in the Dominican Republic and Colombia, while oligotrophic conditions in mid–high latitudes catalysed the widespread dominance by larger benthic Foraminifer assemblages, as observed in Southern Spain.
See also...





Follow Sciency Thoughts on
Facebook.