
Body fossils, otoliths, ichthyoliths, and molecular clocks point to the early Palaeogene as a time of remarkable diversification and innovation among marine fishes. Most of the groups familiar from modern marine ecosystems appeared during this interval, along with their distinctive morphological adaptations. So striking is this pattern that the evolution of marine teleosts after the Eocene has been described by some as ‘mere tinkering’. One of the clearest impacts of the Cretaceous/Palaeogene extinction on teleosts was the extinction of large predatory taxa, which has been implicated in permitting the subsequent diversification of lineages like Mackerel and kin (Scombridae), Barracudas (Sphyraenidae), Billfishes (Xiphiodei) and Jacks (Carangidae) that first appear near the Palaeocene–Eocene boundary. All of these examples belong to a group of Fish called Acanthomorphs, or Spiny-rayed Teleosts, which represent the dominant fish group in marine settings since the beginning of the Cainozoic. However, the hypothesis that opportunity arising from the Cretaceous/Palaeogene extinctions fuelled diversification predicts that other groups may have also experimented with new roles in the early Palaeogene, although this has been little investigated despite its significance for understanding the structure of marine faunas at that time.
In a paper published in the journal Royal Society Open Science on 13 May 2020, Alessio Capobianco of the Department of Earth and Environmental Sciences and Museum of Paleontology at the University of Michigan, Hermione Beckett of the Department of Earth Sciences at the University of Oxford, and Department of Biology at King’s High School for Girls, Etienne Steurbaut of the Royal Belgian Institute of Natural Sciences and the Department of Earth and Environmental Sciences at KU Leuven, Philip Gingerich, also of the Department of Earth and Environmental Sciences and Museum of Paleontology at the University of Michigan, Giorgio Carnevale of the Dipartimento di Scienze della Terra at the Università degli Studi di Torino, and Matt Friedman, again of the Department of Earth and Environmental Sciences and Museum of Paleontology at the University of Michigan, report a new early–middle Eocene clade of large-bodied Clupeiform (Herring and Anchovies) Fish, the anatomy of which has been revealed by micro-computed tomography.
This group, represented by the previously described Clupeopsis straeleni from the Ypresian of Belgium and a new genus and species from the Lutetian of Pakistan, is characterised by remarkable dental specialisations: a single row of enlarged dentary teeth combined with a single massive vomerine fang that extends to the ventral margin of the mandibular symphysis. These fossils force a reconsideration of trophic diversity among marine clupeiforms in the early Palaeogene, which are otherwise represented by small-bodied, probable planktivores apparently similar to the vast majority of living Clupeiforms. More broadly, they point to previously unappreciated trophic innovation in an early Palaeogene marine setting that has not persisted to the modern day.
Clupeopsis straeleni is known from a single specimen, MRHNB IG 8630, in the collection of the Royal Belgian Institute of Natural Sciences, a three-dimensionally preserved individual comprising an almost complete skull plus incomplete body extending to the level of the dorsal fin. The specimen measures 278 mm from the tip of the snout to the broken posterior end of the body.
This specimen comes from the Dubois clay pit, Chièvres in Hainaut, Belgium, 3.7 mabove Ypresian basal gravel. This represents the basal part of the Orchies Clay Member, providing an age constraint of approximately 54.40–54.05 million years. The Orchies Clay Member is interpreted as being deposited in an outer neritic marine setting, but its base appears to represent a shallower facies.
Clupeopsis straeleni is a Clupeiform with triangular-shaped skull in lateral and dorsal views; vomer bearing two large tooth pits; single vomerine fang, extending ventrally beyond the mandibular symphysis when jaws closed and representing the largest tooth; dorsolaterally oriented pre-epiotic fossa; robust and slightly curved toothless maxilla measuring approximately 75% of neurocranium length; straight and robust dentary bearing caniniform teeth throughout its length; largest dentary tooth approximately 20% of length of orbital cavity; round, greatly expanded posterior infraorbital covering most of hyomandibula and quadrate and part of the maxilla; hyomandibula slightly reclined posteriorly; small lateral horizontal lamina of the ectopterygoid underlying the orbit; second basibranchial longest element of the basibranchial series.

Clupeopsis straeleni (MRHNB IG 8630, holotype). Photographs of the holotype in (a) right lateral and (b) dorsal views. Photographs of the cranial region of the holotype in (c) right lateral, (d) left lateral, (e) dorsal and (f) ventral views. Scale bars are 10 mm. Capobianco et al. (2020).
The postcranium and surficial aspects of the skull were described previously. Capobianco et al. focus on details of the cranium revealed by external restudy and micro-computed tomography.
The skull is triangular in dorsal and lateral view. The preorbital region of the neurocranium is slightly longer than the postorbital region. The frontals are the largest bones of the skull roof; they are narrow and elongated, broadest at mid-length and slightly tapering posteriorly. Two pronounced longitudinal crests extend on the dorsolateral side of the frontals. These ridges start above the orbits and continue posteriorly on the supraoccipital, where they join medially to form an occipital crest. The supraorbital canal is partially exposed medial to the longitudinal crest, in a sulcus above the orbit. A large median fontanelle is present between the anterior portion of the frontals. Posteriorly, the frontal forms the anterior and dorsal margins of the temporal foramen, which is also bound by the parietal. The parietals are completely separated from each other along the midline by the frontals anteriorly and by the supraoccipital posteriorly. The sphenotic is broad, concave and laterally projecting. The pterotic forms the postero-lateral corner of the neurocranium and is dorsoventrally compressed. The lateral region of the pterotic houses the majority of the recessus lateralis chamber, an intracranial space where several cephalic sensory canals converge. The pterotic bears four distinct openings for the recessus lateralis. The medial region of the pterotic houses a pterotic bulla that is connected with a larger prootic bulla; together they form part of the otophysic connection. The posterior part of the frontal, the parietal and the pterotic form a prominent posttemporal groove. The pterotic, parietal and epioccipital delimit a relatively large pre-epiotic fossa with a dorsolateral opening slightly smaller than the temporal fenestra.

Cranial anatomy of Clupeopsis straeleni(MRHNB IG 8630, holotype). (a) Rendering and (b) line drawing of skull, excluding the opercular series. (c) Rendering and (d) line drawing of braincase, palate and suspensorium in left lateral view. (e) Renderings of left dentary in lateral (top) and dorsal (bottom) views. ( f ) Rendering of braincase in anterior view, highlighting the vomerine fang. (g) Rendering of braincase in dorsal view, with partial transparency to reveal the prootic and pterotic bullae. Colours indicate different cranial regions: neurocranium (purple), jaws, palatoquadrate and cheek bones (blue), dorsal portion of hyoid arch (cyan), branchial skeleton (chartreuse). Roman numerals in italics refer to synapomorphic characters within Clupeomorpha. Abbreviations: aa, anguloarticular; afn, anterior frontal fontanelle; bpr, prootic bulla; bpt, pterotic bulla; d, dentary; ecp, ectopterygoid; enp, endopterygoid; epo, epioccipital; fp, pre-epiotic fossa; fr, frontal; h, hyomandibula; hp, hyomandibular process; ih, interhyal; io, infraorbital; le, lateral ethmoid; llh, lateral laminar process of hyomandibula; mes, mesethmoid; mtp, metapterygoid; mx, maxilla; ors, orbitosphenoid; pa, parietal; pal, palatine; pas, parasphenoid; pto, pterotic; pts, pterosphenoid; q, quadrate; rap, retroarticular process; rec, recessus lateralis; soc, supraoccipital; sp, sphenotic; tf, temporal foramen; vbe, ventral branchial elements; vf, vomerine fang; vo, vomer. Scale bar is 10 mm. Capobianco et al. (2020).
The dorsoventrally flattened mesethmoid extends a short distance anterior to the vomer. The irregularly shaped lateral ethmoid forms the anterior margin of the orbit. The vomer houses two very large tooth sockets, of which only the right is occupied. It bears a massive fang that projects ventrally beyond the mandibular symphysis. The vomerine fang is laterally compressed and slightly curved posteriorly. The toothless parasphenoid is elongated, dorsoventrally compressed and slightly arched. Below the otic region of the braincase, it divides into two slender branches that extend posteriorly beyond the occipital condyle.
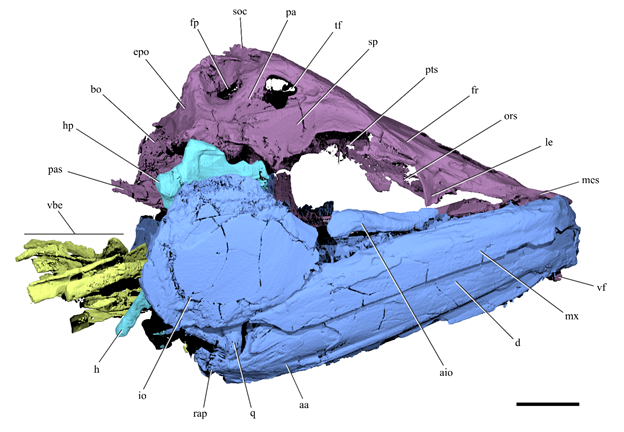
Cranial anatomy of Clupeopsis straeleni. Rendering of Clupeopsis skull in right lateral view, excluding the opercular series. Colours indicate different cranial regions: neurocranium (purple), jaws, palatoquadrate and cheek bones (blue), dorsal portion of hyoid arch (cyan), branchial skeleton (chartreuse). Abbreviations: aa, anguloarticular; aio, anterior infraorbital; bo, basioccipital; d, dentary; epo, epioccipital; fp, pre-epiotic fossa; fr, frontal; h, hyomandibula; hp, hyomandibular process; io, infraorbital; le, lateral ethmoid; mes, mesethmoid; mx, maxilla; ors, orbitosphenoid; pa, parietal; pas, parasphenoid; pts, pterosphenoid; q, quadrate; rap, retroarticular process; soc, supraoccipital; sp, sphenotic; tf, temporal foramen; vbe, ventral branchial elements; vf, vomerine fang. Scale bar is 10 mm. Capobianco et al. (2020).
The jaws extend posteriorly beyond the orbit. The maxilla is toothless, relatively deep and robust. Even though the maxilla is completely devoid of teeth, its ventral margin appears to bear micro-serrations along most of its length.Wewere not able to identify any premaxillae or supramaxillae. Fragments of bone anteromedial to the maxilla might represent parts of the premaxilla. The mandible includes both a dentary and an anguloarticular; the presence of a retroarticular cannot be determined. The dorsal margin of the dentary bears a row of 11–12 caniniform teeth. The first two teeth are smaller than the others and positioned at the anterior tip of the lower jaw. Some teeth are separated from each other by pits. The coronoid process is not strongly developed. A low retroarticular process is present on the anguloarticular.

Neurocranium of Clupeopsis straeleni. Renderings of Clupeopsis neurocranium in (a) dorsal and (b) ventral views. Scleral ring and vomerine fang not shown for clarity. (c) Rendering of †Clupeopsis neurocranium in left lateral view, with partial transparency to reveal the morphology of the recessus lateralis. The recessus lateralis chamber has been digitally filled to highlight its extent and its external openings in the braincase. Abbreviations: acr, accessory temporal sensory canal opening to the recessus lateralis chamber; afn, anterior frontal fontanelle; aor, anterior opening to the recessus lateralis chamber; exo, exoccipital; fp, pre-epiotic fossa; fr, frontal; frco, opening in frontal to supraorbital canal; frlc, longitudinal crest of frontal; le, lateral ethmoid; mes, mesethmoid; mor, middle opening to the recessus lateralis chamber; ors, orbitosphenoid; pa, parietal; pas, parasphenoid; por, posterior opening to the recessus lateralis chamber; ptg, post-temporal groove; pro, prootic; pto, pterotic; pts, pterosphenoid; rec, recessus lateralis chamber; soc, supraoccipital; sp, sphenotic; sr, scleral ring; tf, temporal foramen; vf, vomerine fang; vo, vomer; vots, vomerine tooth socket. Scale bar is 10 mm. Capobianco et al. (2020).
The hyomandibula is directed obliquely, with its distal tip posterior to its articular head. Its ventral arm extends to the level of the quadrate, while its dorsal head bears an extensive lateral laminar process that overlaps the metapterygoid. The interhyal is small and cylindrical. The quadrate bears a large laminar outgrowth at its anterior margin and an anteriorly directed articular condyle at its ventral corner. The quadrate almost completely obscures the large, wedge-shaped symplectic in lateral view. The anterodorsal margin of the metapterygoid bears a deep notch. The ectopterygoid and palatine together form the slender anterior ramus of the palate. Poorly resolved projections on the ventral margin of the palatine might represent teeth. The palatine presents a posteriorly directed flat surface that, together with the lateral ethmoid that directly overlies it, forms the anterior margin of the orbit. The ectopterygoid bears a small lateral horizontal lamina under the orbital region. The endopterygoid is plate-like and lies in a horizontal plane.

Palate and suspensorium of Clupeopsis straeleni. Rendering of left lateral palato-suspensorium of Clupeopsis in medial view. Abbreviations: enp, endopterygoid; ih, interhyal; h, hyomandibula; hp, hyomandibular process; mtp, metapterygoid; pal, palatine; q, quadrate; sym, symplectic. Scale bar is 10 mm. Capobianco et al. (2020).
The gill skeleton is partially articulated, but the identity and morphology of individual elements is difficult to establish due to poor contrast in tomograms. Dorsal and ventral hypohyals are clearly separate and articulate posteriorly with the anterior ceratohyals. There is no trace of a basihyal nor of a basihyal toothplate. The basibranchial toothplate appears to be fused only to the second basibranchial. The second basibranchial is the longest of the basibranchial series.

Lateral horizontal lamina of the ectopterygoid in Clupeopsis and Monosmilus, as seen in cross-sectional tomograms. Transverse plane tomograms of (a) Clupeopsis straeleni and (b) Monosmilus chureloides, highlighting the ectopterygoid(s) in violet and the maxilla(e) in blue. Original tomograms have been mirrored to have left side of the specimens on the left and right side of the specimens on the right. Abbreviations: ecp, ectopterygoid; lle, lateral lamina of the ectopterygoid; mx, maxilla. Scale bars are 2 mm. Capobianco et al. (2020).
Only two infraorbitals are identifiable. The first, representing either the first or second infraorbital, is elongated and slightly curved and lies ventral to the anterior portion of the orbit. The second, which probably represents the third infraorbital based on its position, is rounded and greatly expanded, covering most of the hyomandibula, quadrate and the posterior region of the maxilla. The infraorbital sensory canal extends through the anterior portion of the third infraorbital. Only discontinuous segments of the canal can be identified in the anterior elongated infraorbital. One scleral ossicle can be identified on the left side of the specimen.

Branchial skeleton of Clupeopsis straeleni. Renderings of the gill skeleton of Clupeopsis in dorsal (top) and left lateral (bottom) views. Elements whose identity is uncertain are labeled with a question mark followed by a tentative identification. Abbreviations: bb1, basibranchial 1; bb2, basibranchial 2; bb3, basibranchial 3; bbtp, basibranchial toothplate; cb1, ceratobranchial 1; cb2, ceratobranchial 2; cb3, ceratobranchial 3; cb4, ceratobranchial 4; cb5, ceratobranchial 5; cha, anterior ceratohyal; chp, posterior ceratohyal; dh, dorsal hypohyal; hb1, hypobranchial 1; hb2, hypobranchial 2; hb3, hypobranchial 3; iph2, infrapharyngobranchial 2; iph3, infrapharyngobranchial 3; vh, ventral hypohyal. Scale bar is 10 mm. Capobianco et al. (2020).
While the opercular series is present in the specimen individual bones are difficult to identify in the tomograms due to poor contrast. The preopercle has a gently curved anterior margin, with no clear distinction between dorsal and ventral limbs. The opercle appears to be relatively large, with length roughly equal to its depth.
Secondly, a new species is described and named Monosmilus chureloides. The generic .derives from the combination of the Ancient Greek 'mónos' meaning 'single' and 'smil’e' meaning 'knife', referring to the single massive vomerine fang; the specific name derives from the combination of 'Churel', the name in Urdu of a shapeshifting vampire-like demon with large fangs or tusks, with the suffix '-oides', indicating similarity.
This is described from a single specimen, GSP-UM 37, in the collection of the Geological Survey of Pakistan, in Quetta, Pakistan. Holotype and only known specimen, an incomplete but three-dimensionally preserved skull broken anteriorly at the tip of the snout and posteriorly in advance of the occipital condyle. The specimen measures 104 mm between these broken surfaces. It was collected during a November–December 1977 field season of the Geological Survey of Pakistan and the University of Michigan Museum of Paleontology.

Cranial anatomy of Monosmilus chureloides (GSP-UM 37, holotype). (a) Rendering and (b) line drawing
of specimen in left lateral view. (c) Rendering of basibranchial series with hypohyals (dark green) in left lateral view. (d ) Rendering
of braincase in anterior view, highlighting the vomerine fang. (e) Rendering of right dentary (incomplete) in medial view, with loose
tooth crowns ( probably replacement teeth) highlighted in red. Colours indicate different cranial regions: neurocranium ( purple),
jaws, palatoquadrate and cheek bones (blue), dorsal portion of hyoid arch (cyan), branchial skeleton (chartreuse). Abbreviations: afn, anterior frontal fontanelle; bb1, first basibranchial; bb2, second basibranchial; bb3, third
basibranchial; bbtp, basibranchial toothplate; d, dentary; dh, dorsal hypohyal; f, frontal; ecp, ectopterygoid; enp, endopterygoid;
h, hyomandibula; io, infraorbital; le, lateral ethmoid; mpt, metapterygoid; mx, maxilla; ors, orbitosphenoid; pal, palatine; pas,
parasphenoid; q, quadrate; sp, sphenotic; sr, scleral ring; vf, vomerine fang; vh, ventral hypohyal; vo, vomer. Scale bar is 10 mm. Capobianco et al. (2020).
The specimen was collected from Rakhi Nala (locality RN-4) on the east side of the Sulaiman Range in the Dera Ghazi Khan District of western Punjab Province, Pakistan. The ‘lower chocolate clays’ of the Domanda Formation were deposited in a shallow coastal marine environment. Nannoplankton stratigraphy (zone NP 15) constrains the Domanda Formation to the Lutetian stage/age of the early–middle Eocene.

Monosmilus chureloides, holotype (GSP-UM 37). (a) Simplified map of Pakistan showing the location of the Rakhi Nala locality, where the holotype of Monosmilus has been found. (b) Stratigraphic sequence of Eocene deposits exposed in Rakhi Nala, highlighting the Domanda Formation where the holotype of Monosmilus has been found. For the sequence stratigraphy column, shaded regions represent relative sea level and ‘u’ indicates an unconformity. Photographs of the Monosmilus chureloides holotype in (c) left lateral and (d) right lateral views. Scale bars (a) 200 km; (c), (d) 10 mm. Capobianco et al. (2020).
Monosmilus chureloides is a Clupeiform with a vomer bearing two large tooth pits; single vomerine fang representing the largest tooth; parasphenoid straight in lateral view; orbitosphenoid antero-ventrally contacting the parasphenoid; occipital region posteriorly elongated; robust and straight maxilla lacking teeth; dentary bearing greatly enlarged, postero-medially recurved caniniform teeth throughout its length; largest dentary tooth approximately 70% of length of orbital cavity; bulbous lateral ethmoids; anterior part of palatine triangular in lateral view; horizontal lamina of the ectopterygoid under the orbit and directly overlying the maxilla; third basibranchial longest element of the basibranchial series.
The attribution of two closely related species possibly forming a monophyletic group to two separate genera is subjective. Monosmilus chureloides differs from Clupeopsis straeleni in several morphological features, including: proportionally larger teeth on vomer and dentary; bulbous (rather than irregularly flattened) lateral ethmoid; postorbital region of the neurocranium longer (instead of shorter) than preorbital region; contact between orbitosphenoid and parasphenoid; larger lateral horizontal lamina of the ectopterygoid; longer and broader endopterygoid; third (instead of second) basibranchial as the longest element of the basibranchial series. We consider these differences sufficient to justify the erection of a new genus for Monosmilus chureloides.
The dorsal part of the neurocranium of Monosmilus chureloides is incomplete, with only the eroded mesethmoid, frontals and left sphenotic present in the orbital and ethmoid regions. The posterodorsal part of the neurocranium is missing, along with the occiput.
The narrow, elongate vomer underlies the ethmoid region, and terminates at a narrow point in the orbital region. It bears two large, deep pits, of which only the left is occupied by a massive, laterally compressed fang. The vomerine fang bears a weakly developed keel along its posterior margin. Its distal tip is broken, so its extent is not clear. The lateral ethmoids are large and bulbous and define the anterior margin of the orbits. The parasphenoid is almost straight in lateral view, only curving gently dorsally in the occipital region. It is plank-like, dorsoventrally compressed in the orbital region. Below the otic region it splits into a pair of deep and laterally compressed processes, which extend at least back to the broken posterior margin of the specimen. The foramen for the internal carotid pierces the parasphenoid immediately posterior to the orbital region. The antero-ventrally expanded orbitosphenoid extends to the parasphenoid in the anteriormost part of the orbital region, forming a partial septum. The incomplete sphenotic bears part of the anterior hyomandibular facet. Posterior to this facet, two large foramina pierce the prootic. The more anterodorsal of these is for the hyomandibular branch of cranial nerve VII. Although it is incomplete, the postorbital region of the neurocranium is longer than the preorbital region.

Neurocranium of Monosmilus chureloides. Renderings of Monosmilus neurocranium in (a) dorsal, (b) ventral and (c) left lateral views. Scleral rings and vomerine fang not shown in (a) and (b) for clarity. Abbreviations: afn, anterior frontal fontanelle; fh, foramen for the hyomandibular trunk of cranial nerve VII; fr, frontal; icf, foramen in parasphenoid for internal carotid artery; le, lateral ethmoid; ors, orbitosphenoid; pas, parasphenoid; pro, prootic; sp, sphenotic; sr, scleral ring; vf, vomerine fang; vo, vomer; vots, vomerine tooth socket. Scale bar is 10 mm. Capobianco et al. (2020).
Within the upper jaw, only the maxilla is preserved, albeit incompletely. It is straight, relatively robust and devoid of teeth. The lower jaws are fragmentary, represented by partial dentaries bearing at least eight large, caniniform teeth that are slightly recurved postero-medially. These are generally separated from each other by deep pits. Six to seven loose tooth crowns are preserved parallel to the long axis of the jaw. Some of these occupy pits and appear to be replacement teeth that are not yet connected by bone to the underlying jaw. The external lamina of the dentary partially covers the bases of the teeth.

Cranial anatomy of Monosmilus chureloides. Rendering of Monosmilus skull in right lateral view. Elements whose identity is uncertain are labeled with a question mark followed by a tentative identification. Colors indicate different cranial regions: neurocranium (purple), jaws, palatoquadrate and cheek bones (blue), dorsal portion of hyoid arch (cyan), branchial skeleton (chartreuse). Abbreviations: cb1, ceratobranchial 1; cb2, ceratobranchial 2; cb3, ceratobranchial 3; d, dentary; ecp, ectopterygoid; enp, endopterygoid; f, frontal; h, hyomandibula; le, lateral ethmoid; mtp, metapterygoid; mx, maxilla; pal, palatine; pas, parasphenoid; ors, orbitosphenoid; sr, scleral ring; vf, vomerine fang; vo, vomer. Scale bar is 10 mm. Capobianco et al. (2020).
There is no clear separation between palatine and ectopterygoid, which appear to be toothless with the exception of one or two teeth on the ventral margin of the palatine. The anterior portion of the palatine is triangular in lateral view and is bounded postero-medially by a flat and broad surface, which, together with the posterior surface of the lateral ethmoid, forms the anterior margin of the orbit. In the orbital region, the palatine-ectopterygoid is dorsoventrally compressed and bears a lateral horizontal lamina that overlies the maxilla. The endopterygoid is toothless and large, bearing two transverse ridges that divide its dorsal surface into three concave fossae. The metapterygoid is incomplete posteriorly but has a strongly concave anterior margin. A broken sliver of bone represents all that remains of the quadrate. The anterior extent of the quadrate is constrained by the dentary, and it is apparent that the jaw joint was located far posteriorly. This position, combined with the more anterior location of the small portion of the hyomandibular head that is preserved, indicates that the hyomandibula was strongly inclined posteriorly.
The dorsal and ventral hypohyals articulate posteriorly with the incompletely preserved and platelike anterior ceratohyal. There is no ossified basihyal or basihyal toothplate, but there is a well-developed basibranchial toothplate that is tightly fused to the second and third, but not first, basibranchials. The second basibranchial is longer than the first and bears long flanges that extend ventrally from its lateral surface. The third basibranchial is the longest, with the toothplate covering only its anteriormost portion. Several additional components of the gill skeleton are preserved, including well-developed hypobranchials and ceratobranchials and part of the dorsal gill skeleton, but individual bones are displaced and difficult to identify. Rakers, if present, must have been very small..

Branchial skeleton of Monosmilus chureloides. Renderings of the gill skeleton of Monosmilus in left lateral (top) and right lateral (bottom) views. For paired bones, left-side elements are labeled only in left lateral view, while right-side elements are labeled only in right lateral view. Elements whose identity is uncertain are labeled with a question mark followed by a tentative identification. Abbreviations: bb1, basibranchial 1; bb2, basibranchial 2; bbtp, basibranchial toothplate; cb1, ceratobranchial 1; cb2, ceratobranchial 2; cb3, ceratobranchial 3; cha, anterior ceratohyal; dh, dorsal hypohyal; eb1, epibranchial 1; eb2, epibranchial 2; eb3, epibranchial 3; gf, mineralized gill filaments; hb1; hypobranchial 1; hb2, hypobranchial 2; iph1, infrapharyngobranchial 1; iph2, infrapharyngobranchial 2; vh, ventral hypohyal; uh, urohyal. Scale bar is 10 mm. Capobianco et al. (2020).
Impressions and bone fragments on the outer surface of the specimen represent infraorbitals, but their shape and number cannot be determined. Two scleral ossicles are preserved on the left side and one on the right side of the specimen.
Clupeopsis and Monosmilus are united by a peculiar modification of the vomerine dentition: a pair of laterally adjacent tooth loci, of which only one is occupied at a time by a massively enlarged fang. Expanded vomerine fangs are rare in Teleosts, and to Capobianco et al.'s knowledge, no groups share the asymmetric pattern common to these two Eocene taxa. Instead, other arrangements include a single midline fang (e.g. the Deep-sea Eel, Monognathus, the Pearlfish, Pyramodon), a row of midline fangs (e.g. some Dysommine Cutthroat Eels, some Ophichthid Snake Eels, the Cretaceous Aulopiform, Cimolichthys), paired fangs (e.g. Smelt, Osmerus, some Lophiid Anglerfishes) or multiple longitudinal rows of fangs (e.g. some Serrivomerid Sawtooth Eels and some Ophichthid Snake Eels). Clupeopsis and Monosmilus bear this unusual vomerine dentition in conjunction with a single row of greatly enlarged dentary fangs in association with reduced dentition on the maxilla. In addition to their specialized dentition, both Clupeopsis and Monosmilus share a lateral horizontal lamina of the ectopterygoid that overlies the maxilla.
Evidence that Clupeopsis and Monosmilus are Clupeomorphs, and more specifically Clupeoids (Herring), derives from the better-known Clupeopsis. Capobianco et al. use small Roman numerals refer to some characters. Clupeopsis shows the following Clupeomorph synapomorphies: (i) pterotic and prootic bulla associated with otophysic connection and (ii) a well-defined pre-epiotic fossa. Clupeiform synapomorphies apparent in Clupeopsis are (iii) parietals separated by frontals and supraoccipital and (iv) a recessus lateralis chamber. Finally, Clupeopsis shares with Clupeoidei (v) the absence of a trunk lateral line canal. None of these regions is preserved in Monosmilus. Instead, the placement of Monosmilus within these groups is based on strong evidence that it is closely related to Clupeopsis.
Clupeopsis and Monosmilus share numerous derived characters with Engraulids (Anchovies), but in many cases, these features show a less extreme form of the condition in the Eocene genera than in modern Anchovies: (vi) suspensorium posteriorly inclined, but less than in modern Engraulids (observed in Clupeopsis, inferred in Monosmilus); (vii) mesethmoid projecting in advance of vomers, but less than in modern Engraulids (observed in Clupeopsis, but cannot be checked in Monosmilus); (viii) substantial portion of metapterygoid situated anterodorsal to quadrate (apparent in both genera, but probably linked to suspensorium angle); (ix) ventral limb of hyomandibula meeting the posterior margin of the quadrate (observed in Clupeopsis, but cannot be checked in Monosmilus); and (x) absence of bony basihyal and basihyal toothplate (observed in Monosmilus, probably in Clupeopsis). To these established characters, Capobianco et al. add three observations that support Engraulid affinities. First, both genera share with Engraulids a straight maxilla that contrasts strongly with the curved geometry characteristic of other Clupeomorphs. Second, like Engraulids but unlike other Clupeomorphs, Clupeopsis has a greatly reduced coronoid process (condition in Monosmilus cannot be assessed). Third, Clupeopsis shares with Engraulids an expansion of the lateral laminar process of the hyomandibula that overlaps part of the metapterygoid (condition in Monosmilus cannot be assessed).

Comparative morphology of Sabre-toothed stem-Anchovies and other Clupeiforms. Left lateral views of braincases, palatosuspensoria and basibranchial/basihyal series belonging to representative Clupeiforms. From left to right: Odaxothrissa mento (Clupeidae), Chirocentrus dorab (Chirocentridae), Sabre-toothed stem-Anchovies (braincase and palato-suspensorium of Clupeopsis straeleni, basibranchial series of Monosmilus chureloides), Setipinna (Lycothrissa) crocodilus (Engraulidae). Synapomorphies of successively restrictive Clupeiform sub-clades are indicated in italics Roman numerals and marked over a simplified phylogeny of Clupeoidei, indicating the stem engraulid position of Clupeopsis and Monosmilus and the uncertain relationships at the Clupeoidei base. Colours indicate different sets of cranial bones: neurocranium ( purple), palate (violet), quadrate and symplectic (blue), hyomandibula and interhyal (cyan), basibranchials (chartreuse), basihyal (ochre green, if present), hypohyals (dark green). Renderings not at same scale. Capobianco et al.'s (2020).
Clupeopsis and Monosmilus retain primitive traits that exclude them from the Engraulid crown. These genera lack both fusion between the first basibranchial and the basibranchial toothplate, as well as transverse bony struts enclosing an enlarged supraorbital canal in the frontal. Moreover, Clupeopsis has an anteriorly, rather than posteriorly, directed articular surface of the quadrate (condition in Monosmilus cannot be assessed).
Clupeopsis and Monosmilus are large in comparison to extant Anchovies and indeed Clupeiforms more generally. The incompleteness of the two specimens precludes exact measurements, but estimates can be made based on proportions in living species. Linear regressions of body length with respect to head length in extant clupeiforms yield body lengths of just below half a metre and one metre for Clupeopsis and Monosmilus, respectively. These large sizes in conjunction with well-developed caniniform dentition and slender mandibles consistent with rapid jaw closing suggest a predatory, and probably piscivorous, feeding ecology for Clupeopsis and Monosmilus. The majority of modern Clupeiforms are planktivores. Similar ecologies are inferred for most fossil forms (but see the Early Cretaceous Cynoclupea). However, among extant Clupeiforms, piscivory is the second-most common dietary strategy and appears to have evolved multiple times independently from zooplanktivory. Extant piscivorous taxa have a variety of tooth morphologies, including complete absence of teeth, but all Clupeiforms with caniniform teeth are piscivorous. These include members of Chirocentridae (Chirocentrus), Clupeidae (Odaxothrissa), Pristigasteridae (Chirocentrodon bleekerianus) and Engraulidae (Setipinna (Lycothrissa) crocodilus and Lycengraulis). Among these, only Chirocentrids (Wolf Herring) show a development of the caniniform dentition comparable to that of Monosmilus and Clupeopsis.
The relationships among major Clupeoid lineages are unclear. Genomic-scale data are available for few species, while taxonomically well-sampled molecular phylogenies include only a handful of loci. The placement of Chirocentrids is especially unclear, with this group resolved in a variety of positions by different molecular analyses. A sister-group relationship between Engraulids and Chirocentrids on the basis of shared features in multiple anatomical systems has been hypothesised. Capobianco et al.'s placement of the large-fanged Monosmilus and Clupeopsis as stem Engraulids provides a new perspective bearing on the hypothesised relationship between Chirocentrids and Anchovies. Although these Eocene genera clearly possess many derived features of Engraulids that are absent in chirocentrids, the overall structure of mandibular dentition (in both genera) and braincase (in Clupeopsis; incomplete in Monosmilus) is remarkably similar to that of Chirocentrus. These features could represent generalized conditions of the putative Engraulid/Chirocentrid radiation, subsequently lost in more crownward Engraulids.

Histogram displaying the distribution of body sizes in extant Engraulid (orange) and other Clupeiform (brown) species. Approximate estimates for Clupeopsis straeleni and Monosmilus chureloides were obtained by performing linear regressions of head length vs standard length in extant Chirocentrid and Engraulid specimens. Chirocentrids were chosen because they represent the upper limit of body sizes in extant Clupeiforms and because Clupeopsis likely had similar body proportions (based on what is preserved in the only known specimen for this taxon); Engraulids were chosen because they are the closest living relatives of the two fossil taxa. The head length of Clupeopsis was measured at 75 mm, while the head length of Monosmilus was estimated at 150 mm (based on the incomplete skull representing the only known specimen for this taxon). Head length and standard length measurements for the linear regressions were taken from specimen photographs of Chirocentrids and Engraulids in FishBase. Capobianco et al.'s (2020).
The Early Cretaceous Cynoclupea joins Chirocentrus as another putative relative of the Engraulid clade, which has led to the hypothesis that planktivorous Engraulids are derived from a piscivorous ancestor. This dietary transition had not been inferred elsewhere in Clupeoids, but has clear precedents in other groups of modern (e.g. Polyodon, Scombrine Scombrids) and extinct (e.g. Edentulous, Pachycormiforms) Fish. Capobianco et al.'s interpretation of Monosmilus and Clupeopsis could provide additional support for this inferred trajectory of trophic evolution. Future analyses including morphological data and expanded molecular datasets will represent a vital test of the relationships among modern lineages, hopefully providing a more robust framework for considering the meaning of Clupeopsis and Monosmilus for the evolution of feeding strategies within Clupeiforms.
Independent of their remarkable anatomical specialisations, Clupeopsis and Monosmilus provide a rare and unexpected perspective on the fossil record of total group engraulids. Despite including over 150 extant species that are widely distributed, Engraulids have a very poor fossil record both in absolute terms and in comparison to other Clupeoids. Until the discovery of the early Eocene (Ypresian) crown Engraulid Eoengraulis from Bolca, definitive fossil Anchovies were known only from Miocene or younger deposits. Dated at roughly 5 million years older than Eoengraulis, Clupeopsis is the oldest representative of the Engraulid total group, providing a fossil-based minimum age for the divergence between this radiation and its yet unresolved sister lineage. However, the divergence between Engraulids and other Clupeiforms is likely to substantially predate the age of Clupeopsis, with molecular clock estimates placing the origin of the engraulid total group anywhere from the Early Cretaceous to the earliest Palaeogene. The persistence of fanged stem Engraulids for an interval of at least 8 million years from the early Ypresian to the Lutetian points to the early–middle Eocene as an interval of high engraulid morphological and ecological disparity, with these trophically specialised, early-diverging lineages coexisting with highly nested members of the crown radiation that, like most modern species, were probably planktivorous.
One of the most striking features of turnover among marine Fish associated with the Cretaceous/Palaeogene boundary is the extinction (e.g. Pachycormids, Ichthyodectiforms, Cimolichthyids and some other Aulopiforms) or decimation and marginalisation (e.g. Pachyrhizodontids, Dercetids) of many of the dominant groups of large, predatory Cretaceous Fish, followed by the Palaeogene origins of several independent lineages of predatory Acanthomorphs (e.g. Sphyraenids, Carangids, Xiphioids, Scombrids, Trichiuroids, along with the short-lived Rhamphognathids and Mesogasterids) associated with the broader radiation of Spiny-rayed Fish.
In comparison, the patterns of trophic diversification and environmental shifts among non-Acanthomorph lineages around the Cretaceous/Palaeogene have received less attention. However, there are striking cases of apparently short-lived predatory non-Acanthomorph lineages in shallow marine settings that, like the origin of modern groups of predatory Acanthomorphs, might represent a response to new ecological opportunities in the early Palaeogene. The most conspicuous example is a radiation of Osteoglossids, a group that in the modern day is restricted to freshwater settings. Marine Osteoglossids range in age from early Palaeocene (Danian) to middle Eocene (Lutetian) and include several named genera and unnamed forms. Marine Osteoglossids partially overlapped temporally, and in some cases co-occurred with, an early Eocene (Ypresian) to early Oligocene (Rupelian) shallow-water lineage of large-bodied (about one metre) Paralepidid Aulopiforms (Barracudinas), a group that in the modern day is associated with meso- or bathypelagic settings. These new Palaeogene radiations were joined by a handful of late-surviving examples of predatory marine Neopterygian lineages that arose in the Mesozoic, all of which (e.g. Serrasalmimid Pycnodontiforms, Xidalamine Amiids and possibly Pachyrhizodontids) appear to have gone extinct by the end of the Eocene. The raptorial stem Engraulids Clupeopsis and Monosmilus add to this diverse assortment of early Palaeogene non-Acanthomorph predators in shallow-water settings. It is unclear whether the ecology characterising these genera evolved in the Cretaceous, or if it only arose in the Palaeogene. However, it is apparent that Palaeogene predatory Fish guilds in shallow marine settings were composed of phylogenetically diverse members, paralleling the pattern of numerous now-extinct predatory Mammal and Archosaur lineages (e.g. Creodonts, Mesonychids, Sparassodonts and terrestrial Crocodilians) that coexisted with modern radiations (e.g. Carnivorans) in terrestrial ecosystems at the same time.
See also...






Follow Sciency Thoughts on
Facebook.