
Echidnas (Tachyglossidae) represent only one of two living families of Monotremata, an early-diverging lineage of egg-laying Mammals. Generally classified as fossorial specialists, Echidnas possess robust pectoral girdle and limb bones, a broad manus, and long, spade-like claws. However, within the forelimb they also display many anatomical features reminiscent of earlier Mammalian relatives. These include additional bones within the pectoral girdle (an interclavicle fusedwith the clavicle, a coracoid fused to the scapula, and an epicoracoid), a laterally-facing hemi-sellar glenoid, lack of a scapular spine, and similar general humeral morphology. Although the subject of few biomechanical studies, their unusual anatomy and phylogenetic placement makes musculoskeletal function in Echidnas of particular interest. Not only can they offer an independent case study on how anatomy is shaped by evolutionary adaptation and constraint, but they can also provide insight on the evolution of the Mammalian musculoskeletal system and ancestral Mammalian function. Echidnas have a characteristic sprawling gait, with rolling of the trunk and a slow, pace-like walk. Walking kinematics show long-axis rotation to be the principal motion of the humerus, unlike most Therian Mammals where the predominant motion is flexion-extension within the parasagittal plane, but likely similar to some earlier relatives of Mammals (e.g., the non-Mammalian Pelycosaurs and Cynodonts). Moment arm estimations based on muscle attachment sites likewise suggest that the shoulder is optimized for internal rotation and adduction. There appears to be little intrinsic muscular stabilization of the pectoral girdle and shoulder joints, due to the robust bony and articular morphology in Monotremes. However, it is unknown whether the muscles themselves are structurally specialised for the Echidna’s lifestyle and locomotion.
A primary determinant of muscle function is muscle architecture, or the arrangement of fibers within muscle. Fiber length is linked to muscle shortening ability and velocity, while muscle physiological cross-sectional area determines how much force a muscle can generate. Gross muscle anatomy has been described qualitatively for the Echidna, but muscle architecture remains unexplored in Monotremes. Studies on forelimb muscle architecture of other Mammals demonstrate how certain muscles have become specialized for different lifestyles and locomotion. For example, the physiological cross-sectional areas of the teres major muscle and the subscapularis muscle in the fossorial Mole, Scalopus aquaticus, are disproportionately large compared to body mass, to produce the forceful humeral abduction and internal rotation necessary for their unusual digging style. The physiological cross-sectional area of the subscapularis muscle is likewise large in the fossorial Badger, Taxidea taxus, with short fibers suggesting this muscle is specialised for stabilisation of the humerus while digging. The physiological cross-sectional area of the subscapularis muslce is also large in the arboreal Pine Marten, Martes martes, to counteract laterally-directed reaction forces in the limb during climbing.
In a paper published in the Journal of Mammalian Evolution on 14 March 2020, Sophie Regnault and Philip Fahn-Lai of the Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology at Harvard University, Rachel Norris of the School of Animal and Veterinary Science at the University of Adelaide, and Stephanie Pierce, also of the Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology at Harvard University, document the shoulder and proximal forelimb muscles and architectural parameters of the Short-beaked Echidna, Tachyglossus aculeatus, providing a detailed examination of muscle form and function in Monotremes.
Muscle data from the Echidna are further compared to other fossorial and non-fossorial Mammals, as well as sprawling non-Mammals, to determine musculoskeletal specialisations associated with the Echidna’s digging lifestyle and sprawling gait. Based on prior work Regnault et al. predicted that the Echidna would show high physiological cross-sectional areas for humeral adductors such as the pectoralis muscle (important for supporting sprawling Animals during stance) and internal rotators such as the teres major muscle (important for the highly specialised fossorial Mole). Regnault et al. also hypothesised convergent specialisations in homologous muscles with fossorial Therians (e.g. the subscapularis muscle). However, as many muscles differ in position, attachments, and inferred function between Echidnas and Therian Mammals, Regnault et al. further anticipated other muscles to reveal differences reflective of the Echidna’s mixture of plesiomorphic and derived anatomical features.
Four adult Short-beaked Echidnas, Tachyglossus aculeatus, were used in this study. One specimen was contrast-stained to obtain a 3D overview of whole muscle anatomy, and three specimens were dissected for architectural properties. The specimens were collected in South Australia, and provided by the University of Adelaide. Cause of death in all cases was suspected impact with a vehicle. The specimens were intact with no grossly observable injuries to the pectoral girdle/forelimb. All specimens had been collected an unknown time after death and stored frozen at −18°C.
The mean architectural parameters and physiological cross-sectional area were calculated for each Animal’s right and left sides. These values were then normalised against each Animal’s body mass (in grams). A mean, body-mass normalised value was then calculated for each muscle. Body mass-normalised parameters allowed broad comparisons to be made between species of different size. Finally, these architectural parameters were also scaled to an Echidna of body mass 3.31 kg, to contextualise architecture in real-world terms and match the mass of the contrast-stained specimen displayed in the gross musculoskeletal anatomy.

Body mass-normalized values of physiological cross-sectional area and fascicle length were compared to normalised muscle architecture parameters calculated from other published Mammal species: the fossorial Mole, Scalopus aquaticus, the fossorial Badger, Taxidea taxus, the cursorial Hare, Lepus europaeus, the arboreal Pine Marten, Martes martes, and the terrestrial Opossum, Didelphis virginiana. Additionally, parameters were compared with two sprawling non-Mammals: the Tegu Lizard, Salvator merianae, and the Alligator, Alligator mississipiensis. Physiological cross-sectional area PCSA was recalculated from the muscle belly masses, pennation angles, and muscle fascicle lengths reported in previous studies, to ensure the same method and scaling was applied to all species. The mean value of each parameter was used from each Animal, apart from the Alligator, where data from an individual of similar mass to the 3D modeled Echidna were used. Muscle architecture parameters were scaled by body mass reported in each study.
The R package ‘dispRity’ was used to compare variance in the mean body mass-normalised architectural parameters (fascicle length and physiological cross-sectional area) of the different species. The disparity metric is equivalent to the mean Euclidean distance between the observed muscle points and a ‘centroid’ (or mean point) on an x-y plot of physiological cross-sectional area vs. fascicle length. To capture the spread as well as the disparity of each species’ muscle architecture, the interquartile range of observed muscle point distances to the centroid were also reported. The disparity metric is calculated as disparity between the single value (e.g. mean body-mass normalised of physiological cross-sectional area/fascicle length) for each muscle of a species i.e. disparity between muscles.
Differentiation of individual muscles via contrast-staining was generally good, despite relatively less diffusion of contrast agent to some very deep parts of the pectoral girdle muscles (e.g., the origin of the subcoracoideus muscle). For ease of description, muscles are classified here into functional groups inferred from estimates of glenohumeral muscle moment arms from a previous study, but note that many muscles cross additional joints and/or have multiple inferred actions. As such, muscle functional group here is classified only according to the largest moment arm produced by that muscle at the glenohumeral joint.
The humeral internal rotators originate and insert near one another. The latissimus dorsi muslce (also called the latissimus spinalis muscle) has a muscular origin from the caudal scapula and vertebral column, with an aponeurotic extension caudally along the vertebral column, and inserts on the edge of the distal humeral entepicondyle. The teres major muscle arises alongside the scapular origin of the latissimus dorsi muscle, and inserts proximally on the crest of the lesser tubercle of the humerus. The subscapularis muscle originates below these, in the subscapular fossa on the external caudal aspect of the scapula, and inserts on the lesser tubercle of the humerus. The teres minor muscle originates above the glenoid fossa, and also inserts on the lesser tubercle. Together, the humeral internal rotator group had the greatest combined muscle mass, principally through the bulky latissimus dorsi and subscapularis muscles.

The humeral external rotators, supraspinatus and infraspinatus muscles, originate cranially on the scapula. The infraspinatus muscle originates from the external aspect, in the infraspinous fossa (separated from the subscapular fossa by the crest of triceps longus muscle), while the supraspinatus muscle originates from the internal scapular surface. Both insert on the greater tubercle of the humerus. The The clavodeltoideus muscle also acts principally to externally rotate and extend the humerus; this muscle originates along the length of the clavicle’s ventral surface, and inserts below the greater tubercle on the cranial aspect of the humerus.

Of the humeral abductors, the acromiodeltoideus muscle originates from the acromion and inserts on a small area on the dorsal aspect of the proximal humerus. This muscle appears to have a split or subdivision in the segmented scan and in dissected specimens. The spinodeltoideus muscle originates on the dorsal edge of the scapula and, enveloped within the clavodeltoideus muscle, inserts near the acromiodeltoideus muscle on the humerus. The coracobrachialis muscle is divided into a long and short head, which originate on the ventral aspect of the coracoid and insert along the ventral aspect of the humerus. The scapular heads of the triceps brachii longus muscles originate from a crest between the infraspinous and subscapular fossae on the external scapula. These two heads are joined by the accessory and lateral heads originating from the caudal aspect of the proximal humerus and the medial head originating from the distal dorsal surface of the humerus. All five heads of the triceps brachii muscle insert on the olecranon process of the ulna.
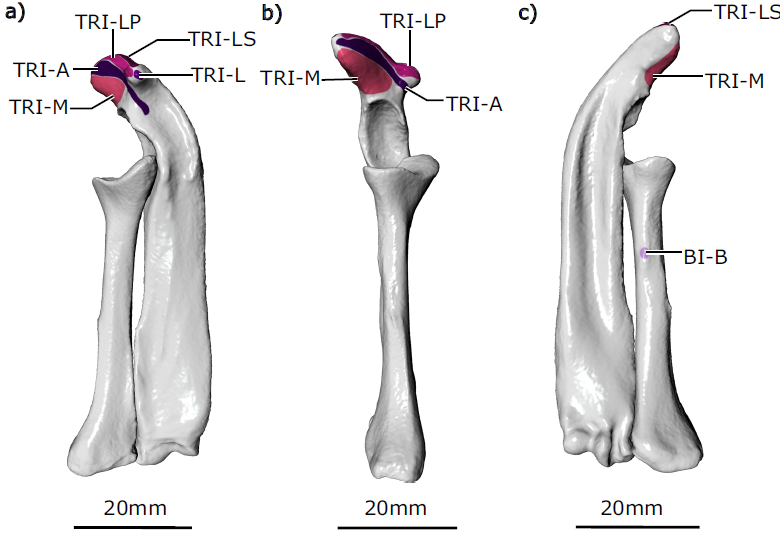
The supracoracoideus muscle and both heads of the biceps brachii muscle act to adduct the humerus; all originate from the ventral epicoracoid and coracoid. The biceps brachii brevis muscle inserts on a small crest on the caudal/ventral aspect of the radius. The biceps brachii longus muscle possesses a longer tendinous insertion and was difficult to trace precisely on the contrast-stained scans, but from dissections appeared to insert on the ulna. The supracoracoideus muscle inserts on the ventral aspect of the humerus’ greater tubercle.
The pectoralis muscle is usually considered a humeral adductor, although in the echidna its moment arms suggest it is capable of producing other equally forceful actions as a humeral internal rotator and extensor. The pectoralis muscle is not subdivided; it originates along the presternum (or manubrium) and three sternal elements, and inserts next to the supracoracoideus muscle on the crest of the greater tubercle of the humerus.
The subcoracoideus muscle appears to be the only muscle whose principal action is humeral flexion. This muscle originates on the internal surface of the epicoracoid and inserts on the caudal humerus, along the lesser tubercle and crest of the lesser tubercle. None of the muscles examined here appear to act principally as humeral extensors (based on relative size of their moment arms), though several are capable of producing humeral extension alongside other movements, e.g. the pectoralis muscle and clavodeltoideus muscle.
Many of the muscles crossing the shoulder generally had little to no external tendon, attaching directly to bone or via short internal tendons or superficial aponeuroses. However, a few muscles did have an appreciable tendon, including the spinodeltoideus muscle (20 mm), the biceps brachii longus and brevis muscles (16 and 12 mm), and the coracobrachialis longus muscle (7 mm).

Many muscles appeared to have fascicles oriented parallel to the muscle’s long axis, with fascicles running along much of the muscle’s length. On closer examination, some of these muscles possessed fascicles inserting directly onto bone, but other fascicles elsewhere within the same muscle inserted at a slight angle onto short internal tendons or superficial aponeuroses (e.g. the pectoralis and latissimus dorsi muscles). Mean pennation angles from these muscles, calculated from angle measurements made at three different muscle regions, ranged from 2 to 13°. Other muscles were clearly parallel-fibered throughout (the acromiodeltoideus, biceps brachii longus, coracobrachialis brevis, supracoracoideus, m. teres minor, and triceps medialis muscles), with pennation angles of 0°. Finally, a third of the examined muscles were more clearly pennate, with mean pennation angles between 15 and 22° (the coracobrachialis longus, infraspinatus, spinodeltoideus, subscapularis, supraspinatus, teres major, and triceps brachii accessorius muscles).
Many of the muscles cluster around similar fascicle lengths, between approximately 20–45 mm, though the latissimus dorsi muscle can be clearly seen to possess the longest mean fascicle length at 78 mm. The long, near parallel structure of fascicles in many of the Echidna’s shoulder muscles, described above, is reflected in calculated ratios of fascicle length to muscle belly length; for all muscles examined, fascicle lengths measured between 52 and 88% of total muscle belly length. Compared to fascicle length, the spread of physiological cross-sectional areas appears more diverse. The muscles with the largest physiological cross-sectional areas were generally those with the largest masses (the latissimus dorsi, the subscapularis, the combined heads of the triceps brachii, and the combined heads of mcoracobrachialis muscle), though many of these muscles had some degree of fascicle pennation that would also have contributed toward increasing physiological cross-sectional area.
In general, the Echidna shoulder muscles exhibited much less architectural variation compared with the other Mammals. The Echidna has the lowest observed disparity (0.72), followed by the Tegu (0.97), Alligator (1.07), Badger (1.39), Pine Marten (1.47), Opossum (1.63), Hare (1.97), and Mole (2.38).
Relative to each muscle’s length and mass, the Echidna possesses muscles with relatively long fascicles and small-to-intermediate physiological cross-sectional areas. These values are similar to the range exhibited by Tegu muscles (fascicles between 54 and 93%), and many of the Mole muscles (fascicles between 63 and 100%, excluding the supraspinatus and infraspinatus muscles). In contrast to the Echidna, the Alligator, Opossum, Hare, and Pine Marten muscles have more variable relative muscle fascicle lengths and physiological cross-sectional areas.
The main aim of Regnault et al.'s study was to characterise the architecture of the shoulder and proximal forelimb muscles in the Monotreme Echidna, to better understand forelimb function and evolution in this relatively under-studied but phylogenetically important group of Animals. Regnault et al. anticipated that the Echidna would show muscle architectural specialisations related to its lifestyle and locomotion. Namely, they hypothesised increased force-generating capacity (via large physiological cross-sectional area) for humeral internal rotators and adductors, which have been identified as important in the fossorial sprawling Echidna in a previous study, and would be convergent with other fossorial Mammals (e.g. Moles) and sprawling non-Mammals (e.g. Crocodilians).
Both Echidnas (Monotremata: Tachyglossidae) and Moles (Placentalia: Talpidae) are thought to be humeral rotation diggers. Although separated by long evolutionary distances they share some anatomical similarities, including a robust humerus with convergent ‘hourglass’ morphology, bulky pectoral girdle, and broad spade-like manus with restricted joint mobility. Many muscles appear to act similarly in the Mole and Echidna, particularly to produce internal humeral rotation. Counter to their expectations, Regnault et al. found that Echidna body mass-normalised muscle architecture parameters were not appreciably convergent to the mole or other fossorial Mammals examined. Unlike the Mole, teres major muscle in the Echidna appeared unremarkable in terms of physiological cross-sectional area. The subscapularis muscle was relatively large and had a larger physiological cross-sectional area than the other Echidna shoulder muscles, but even this muscle did not show the same extreme specialisation relative to body mass as in the Mole or Badger. The pectoralis was another muscle Ragnault et al. anticipated might show architectural specialisation, due to its importance as a humeral adductor in sprawling animals. However, it also did not have a large physiological cross-sectional area or long muscle fascicles relative to other muscles in the Echidna shoulder nor when compared with the pectoral muscle(s) of other sprawling and upright Animals.
In fact, the Echidna’s shoulder and proximal forelimb muscle architecture appears fairly unspecialised. Physiological cross-sectional area and fascicle length values showed little spread, and measures of disparity placed the Echidna lowest amongst all the Animals compared by Regnauly et al., closest to the Tegu Lizard than to any of the other Mammals. Of all the Echidna muscles examined, the latissimus dorsi muscle was probably the most divergent, with the longest mean fascicle length and one of the largest physiological cross-sectional areas. This is likely linked to the muscle’s overall anatomy: the latissimus dorsi muscle inserts very distally on the humerus in Monotremes compared with other Animals. Even so, its mean fascicle length does not reach the same extremes as the latissimus dorsi muscle in the Hare, Opossum, or Mole, and neither is its physiological cross-sectional area especially notable when considered against the range of muscle physiological cross-sectional areas exhibited by other Animals. In contrast to the Echidna, all other Mammals examined by Regnault et al. showed much more variation in shoulder muscle physiological cross-sectional areas and fascicle lengths, with the Mole exhibiting the greatest disparity. The muscle parameters used in the calculation of disparity are scaled by body mass, so in part, increased disparity (e.g. Mole, Hare) can be a reflection of an increase in the proportion of the measured forelimb muscle masses (5.4% and 3.4% of body mass in the Mole and Hare, respectively, compared to 1.8% in the Echidna). In these cases, increased disparity results from some forelimb muscles maintaining similar relative mass, while there are dramatic increases in masses of other specific and apparently functionally-relevant muscles (e.g. the teres major and pectoralis muscles in the Mole, and the triceps brachii and pectoralis muscles in the Hare). The latter appear to be driving the relative increase in overall forelimb muscle mass (the two muscles in each example accounting for over 55% of measured forelimb muscle mass for those species).
When architectural parameters are expressed relative to each muscle’s size, another pattern then emerges. In terms of muscle architecture relative to muscle size, the Echidna (and Tegu) again shows little variation. Moles are more variable; however, this appears to be mostly driven through modulation of muscle size. Once muscle size is accounted for, many Mole muscles have similar relative fascicle lengths and physiological cross-sectional areas as in the Echidna. Regnault et al. show that pennation angle and fascicle length become the determinants of physiological cross-sectional area/muscle mass, and of these, fascicle length appears to be the primary source of diversity. Muscle pennation can help to pack in more short fibers/fascicles, but does not appear a prerequisite for achieving high physiological cross-sectional area/muscle mass, e.g. the coracobrachialis muscle in the Alligator has one of the highest physiological cross-sectional area/muscle mass and no pennation. The Echidna shows only a 3.5-fold difference between its shortest and longest fascicle length; Tegus are similar at 3.6, and moles 5.3 (when excluding the supraspinatus and infraspinatus muscles). In contrast, the other Therian Mammals and the Alligator show much greater variation in fascicle length (up to 17.5 times in the Hare), and consequently much more variation in both fascicle length/muscle length and physiological cross-sectional area/muscle mass.
Why are the Echidna shoulder muscles so architecturally similar? Perhaps muscle specialisations are unnecessary. Muscles with short fascicles in highly pennate arrangements, with large resultant physiological cross-sectional areas, are capable of generating large forces but do not shorten much; these muscles are inferred to have specialised roles as joint stabilisers. Examples include the supraspinatus and infraspinatus muscles in Therian Mammals. These stabilising muscles are important in Therians, to compensate for their highly mobile pectoral girdle and glenohumeral joints. However, the Echidna (like earlier Mammaliaforms) has less mobility at the scapula and glenohumeral joint; the robust and interlocking bones and joint morphologies of the Echidna’s pectoral girdle and forelimb may explain why its muscles have not acquired ‘stabiliser’ muscle architecture. Relative to muscle size, the Echidna’s muscles instead have long fascicles and small-to-moderate physiological cross-sectional areas (similar to the Tegu Lizard). Such muscles are capable of relatively greater and more rapid shortening, and are suited for force production over a wide range of the muscle’s range of motion. The Echidna’s low architectural diversity can thus be interpreted as a lack of muscle specialisation into distinct roles (e.g., joint stabilising, or high force production over narrow working ranges for specific movements), and/or a generalised need for muscles with wide working ranges (e.g. potentially as an adaptation for fossoriality).
It is notable that muscle architectural parameters in the Echidna appear to be broadly conservative. Extant Mammals, and particularly Eutherians (Placentals), exploit a huge variety of ecological niches and locomotory strategies, underpinned by modifications of their forelimb into fins, wings, and other specialised structures. The wide variety of physiological cross-sectional areas and fascicle lengths exhibited by the Eutherian shoulder muscles sampled thus far suggests that the remarkable versatility of the Placental forelimb extends to the level of muscle architecture. Metatherians (Marsupials) appear more constrained in forelimb versatility, perhaps because their forelimb structure is linked to their reproductive strategy of climbing to the pouch after birth. Although Regnault et al. examined data from only one Marsupial in their study (the Opossum), its muscle architecture parameters are less diverse than the Eutherian Mammals, and so appear to reflect the generalised constraint on Marsupial forelimb anatomy. Although Monotremes possess a mosaic of plesiomorphic and more derived forelimb anatomical features, no developmental constraints of the forelimb have been identified comparable to Marsupials, and so Monotremes might be expected to exhibit more muscle architectural diversity than the Echidna does.
If the Echidna’s architecture does indeed reflect lack of muscle specialisation into distinct roles (rather than reflecting a fossorial lifestyle), one possibility may be that diversification of Mammalian muscle architecture occurred in the lineage leading to stem Therians or Eutherians, after their divergence from Monotremes, alongside increased diversification of the forelimb more generally. Previous studies have offered evidence that the architecture of many homologous shoulder muscles is statistically similar between Tegu Lizards and Opossums. They interpreted the surprising similarity in functional characteristics as either conservation from the Amniote state or convergence towards a small-bodied ecological generalist phenotype, and link differences to a postural shift from‘sprawling’ to ‘parasagittal’ limb posture and movement in Therians. Though Reghault et al. do not directly compare homologous muscles in their study, they note that the Echidna, phylogenetically bracketed by those two species, also appears to support the notion of conserved muscle architecture in terms of overall diversity. Rehnault et al. found the spread of Echidna muscle architecture values closest to the Tegu, which might be expected given that Monotremes diverged from other Mammals prior to the hypothesised acquisition of parasagittal posture.
In conclusion, Regnault et al. find that the muscle architecture of the Echidna pectoral girdle and proximal forelimb is functionally conservative. Muscle physiological cross-sectional areas and fascicle lengths do not appear modified for specific roles (e.g., stabilisation) to the same extent as in other fossorial Mammals, or indeed any of the other Animals sampled apart from the Tegu. Rather, all the Echidna’s muscles show similar relative architectural parameters, suited for low-to-moderate force production over a wide working muscle range. Regnault et al.'s species comparisons using existing literature data also suggest that the increase in diversity in Eutherian forelimb structure and function may be mirrored by increasing disparity of muscle architecture in these species, though more data are needed to test this. Diversity in muscle architecture in the studied species appears to be primarily driven through modulation of effective fascicle length (and in some cases, relative muscle size). Muscle architectural studies such as ours are therefore of key importance, not only for better understanding forelimb function in extant Animals like the Echidna and other Mammals, but also for their contribution to reconstructing function in extinct taxa.
See also...






Follow Sciency Thoughts on Facebook.
Follow Sciency Thoughts on Twitter.