
In non-Mammalian vertebrates, the jaw joint is formed between the quadrate (or palatoquadrate) of the upper jaw and the articular part of Meckel’s cartilage, a rod of cartilage that runs through the lower jaw. This is known as the primary jaw joint. In Mammals, this function is carried out by a new joint between the dentary and squamosal bones, known as the temporomandibular joint or TMJ in Humans, and is referred to as the secondary jaw joint. In mammals, the bones of the original primary jaw joint have been incorporated into the ear and play a role in hearing. In addition to forming a joint with the articular as part of the primary jaw joint, the Amniote quadrate also articulates with the cranial base. During the evolutionary transition that gave rise to Mammals, the connection between the quadrate and the cranial base simplified. The robust quadrate of Reptiles moved from being attached to up to five separate skeletal elements, able to bear the mechanical force of feeding, to become the diminutive Mammalian incus, suspended by a ligament from a single cranial base bone, the petrosal, in an air-filled cavity allowing sound transmission. At the same time, the Meckel’s cartilage lost its permanent nature, separating the incus and neighbouring malleus from the rest of the jaw in adults. Early Mammal-like Reptiles had a permanent Meckel’s cartilage and joints between the quadrate and articular, and posteriorly between the quadrate and cranial base, similar to extant Reptiles. In Mammaliaforms, such as Morganucodon, both a primary quadrate-articular and a secondary dentary squamosal joint were present, in addition to a joint between the quadrate (incus) and the paraoccipital process of the petrosal. This petrosal and incus joint precedes detachment of the middle ear from Meckel’s cartilage in Mammal evolution. A connection between the future middle ear bones and the cranial base is therefore a feature of fossil Mammaliaforms. In extant Mammals, the proposed homologue of the paraoccipital process is the crista parotica, which forms as a cartilaginous spur off the petrosal and is derived from neural crest cells, distinct to the rest of the petrosal and otic capsule, which are mesodermally derived. Modern Mammals have separated the middle ear from the jaw in adults, and the ossicles (malleus, incus and stapes) are now suspended by ligaments from the cranial base to allow free vibration during sound transmission from the ear drum to the inner ear. Palaeontological evidence indicates that the evolution of the definitive Mammalian middle ear occurred at least twice, once in the lineage that gave rise to Monotremes and once in the Therian (Marsupial and Eutherian) Mammals, while new developmental data suggests that the two groups of Therian Mammals may have each independently acquired the definitive Mammalian middle ear (it has become common to use the term Eutherian Mammals rather than Placental Mammals, as Marsupials have a yolk-sac placenta).
Marsupials and Monotremes, exhibit extreme altriciality, greater than is seen in any Eutherian. This has profound consequences for early feeding as the bones that form the Mammalian jaw joint, the dentary and squamosal, have not fully ossified by the time of birth/hatching. The dentary-squamosal joint forms prior to birth in Eutherian Mammals, and begins to function in the embryo. In the Mouse, gestation is approximately 20 days, with breakdown of Meckel’s cartilage, to separate the lower jaw from the ear bones, following during early postnatal stages. In contrast, the Opossum, Monodelphis, has a short gestation of just 13 days, and is born before development of the dentary-squamosal articulation, which forms between 14 and 20 days after birth. Monotremes hatch out of the egg after 10 days post-oviposition. The formation of the dentary-squamosal joint in Monotremes has recently been followed and shown to form from 10 days after hatching in the Platypus. Breakdown of Meckel’s cartilage in both Marsupials and Monotremes occurs relatively late postnatally, with a robust Meckel’s still evident in nest young Platypuses. There is, therefore, a significant gap between birth and the advent of a functional Mammalian jaw joint in both Marsupials and Monotremes.
The feeding strategies of new-born Mammals vary in extant members of each group of Mammals. Compared to Eutherian Mammals, Marsupials rely on placental support for a relatively short period of time and consequently receive the nutrition required for their development via a lengthy and sophisticated lactation. During their early postnatal life Marsupials attach to the mother’s teat and use the comparatively early developed tongue musculature to suck. In the Grey Short-tailed Opossum, Monodelphis domestica, pups are born after 13 days of embryonic development, which is followed by around 14 days permanently attached to the mother’s teat, after which they detach intermittently from the mother but continue to suckle. Weaning occurs around postnatal day 60. In contrast, young Monotremes do not obtain milk in quite the same way as Therian Mammals due to the absence of teats in the mother. Instead young Monotremes suck up milk vigorously from the flattened but protuberant nipple-like areola on the mother’s abdomen.In the case of Echidnas, these areolae are within the pouch.
Given the lack of a jaw joint at birth, it has been proposed that Marsupials and Monotremes use the connection between the middle ear bones and cranial base to permit feeding prior to the formation of the articulation between the dentary and squamosal and cavitation of the middle ear.
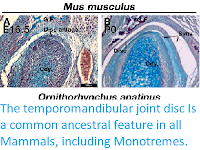
In a paper published in the journal eLife on 30 June 2020, Neal Anthwal of the Centre for Craniofacial and Regenerative Biology at King’s College London, Jane Fenelon of the School of BioSciences at the University of Melbourne, Stephen Johnston of the School of Agriculture and Food Sciences at the University of Queensland, Marilyn Renfree, also of the School of BioSciences at the University of Melbourne, and Abigail Tucker, also of the Centre for Craniofacial and Regenerative Biology at King’s College London, present the results of a study which analysed the articulations that link the lower jaw to the cranial base (cranio-mandibular joints) in Monotremes (Platypus, Ornithorhyncus anatinus, and Short-beaked Echidna, Tachyglossus aculeatus) as they develop from hatching, and compare them to a Marsupial (Grey Short-tailed Opossum, Monodelphis domestica), and a Eutherian (Mouse, Mus musculus), with additional comparison to the Gecko, Guinea Pig and Bat.

Anthwal et al. show that in early post-hatching life the Monotreme incus and cranial base fuse, and later form an articulation, creating a double cranio-mandibular articulation, similar to the jaw anatomy of fossil Mammal-like Reptiles. This close association of the incus and cranial base is also observed at embryonic stages in Eutherians and is reflected in Mouse cell lineage studies. In contrast, Opossums at birth utilise a cushion of extra-cellular matrix-rich mesenchyme in between the incus and petrosal to provide an articulation point. Marsupials and Monotremes, therefore, have different strategies for coping with an early birth. Anthwal et al.'s research suggests that the incus retains a transient lower jaw support role across extant Mammal groups but at differentstages of pre and postnatal development.
It has been suggested that the joint between the malleus and incus might act as the jaw joint early on in Marsupial postnatal development, thereby recapitulating the Reptilian function of these bones in Mammals. Alternatively, it has been suggested that the actual articulation point in Marsupials is between the incus and the cranial base. Less information is available regarding Monotreme development, however, the incus has been described as being in cartilaginous connection with the cranial base during early post hatching development. The development of the malleus and incus, and incus and cranial base, was therefore investigated across the three groups of Mammals, with the Gecko as an outgroup.
In the Ocelot Gecko, Paroedura picta, the quadrate and articular (the homologous elements to the incus and malleus respectively in non-Mammal Amniotes) form a clear synovial joint in the embryo at mid-gestation. In Mice, Mus musculus, the malleus and incus are initially formed from a single cartilaginous condensation that separates, by the formation of a joint, at embryonic day 15.5. At birth, therefore, the incus and malleus are evident as distinct cartilages. In Monodelphis domestica, the malleus and incus are still connected at birth at the dorsal end by a ridge of cartilage. Anthwal et al. observed a similar connection between the malleus and incus in the Echidna, Tachyglossus aculeatus, just after birth. Like the Opossum, the middle ear ossicles were fused dorsally, indicating that they function as a unit. These findings demonstrate that, like Opossums, Monotremes do not use the primary jaw joint as the craniomandibular articulation before the development of the dentary-squamosal joint.

Anthwal et al. therefore investigated the relationship between the incus and the petrosal in the cranial base in Mice, Opossums, Platypus and Echidna, comparing the interaction to the developing joint between the quadrate and opisthotic in embryonic Geckos. In many Reptiles, as shown in the Gecko, the quadrate (incus homologue) forms a synovial joint with the opisthotic (also known as the otoccipital) in the cranial base during embryonic development. The opisthotic/otoccipital is architecturally equivalent to the petrosal of Mammals. In Mice, the crus breve (short process) of the incus nestled in a fossa created by the crista parotica of the petrosal, but was separated by a region of mesenchymal cells, highlighting the lack of a clear articulation point between the two elements. The incus at birth, therefore only articulated with the adjacent middle ear bones, the malleus and stapes. Similar to the Mouse, the crus breve in neonatal opossums, fitted into a fossa created by the crista parotica, but abutted the petrosal on the inferior aspect of the crista parotica. The incus and petrosal were therefore positioned much closer than in the Mouse.
The relationship between the incus and crista parotica in the two Monotreme species was significantly different from the Therian Mammals. In both Platypus, Ornithorhynchus anatinus, and Echidna, Tachyglossus aculeatus, the incus appeared to be fused with the crista parotica at birth. The lower jaw, via Meckel’s cartilage, would therefore be physically connected to the upper jaw, via the incus at this timepoint. The relatively small size of the incus in both monotremes is striking, as is the extended and tapered crus breve of the incus in the Opossum.
To investigate the monotreme relationship between the incus and crista parotica further Anthwal et al. followed development of these two cartilages from birth to functional use of the dentary-squamosal joint, but before complete cavitation of the middle ear space. Due to the scarcity of available specimens very little is known about Monotreme ear and jaw development. In adult Platypuses, the incus appears in contact with the crista parotica, forming a fibrous articulation. Similarly, in the adult Echidna, the incus has been described as tightly attached to the petrosal.
At 2 days and 6.5 days the Platypus incus was fused to the crista parotica by immature chondrocytes. Between 10 days and 30 days the connection was difficult to make out, with the two cartilages almost completely integrated together. Strikingly, by 80 days, when the dentary-squamosal joint would have started to become functional, the incus and crista parotica were no longer fused, with the two distinct cartilages abutting each other. At this stage, in contrast to the other stages investigated, the ear ossicles and petrosal had begun to ossify. However, the regions forming the malleus-incus joint, and the incus-petrosal articulation remained cartilaginous. A cartilaginous articular surface between the incus and petrosal was maintained at 120 days, a period when the young would have started to leave the burrow. A similar move from early fusion, to articulation was observed in the Echidna. No evidence of a synovial capsule, however, was identified at any stage.

The fusion of the incus and crista parotica coincides with the period when the young would have been feeding from milk, while the move to an articulation was associated with periods when the dentary-squamosal was fully formed and functional. After separation of the incus and petrosal, there was a period where two cranial-mandible articulations were evident in the Platypus, between Meckel’s cartilage and the petrosal, via the malleus and incus, and between the dentary and squamosal.
Middle ear cavitation occurred very late in the monotreme specimens analysed, with only the 120 day Platypus showing partial cavitation around the hypotympanum, but this did not extend upwards to where the ossicles are housed. Hearing, thus, must be a very late developing sense in the Platypus.
Limited expression analysis has been performed in monotremes, with no previous expression data performed in the ear or jaw during development. In order to further understand the change in the relationship between the incus and petrosal, immunohistochemistry staining was carried out in Echidna samples 0 and 3 days post hatching.
In the fused incus-petrosal region of 0-day-old Echidna, the expression of both a master regulator of cartilage development, Sox9, and a principal component of cartilage extra cellular matrix, Collagen Type 2, were continuous between the incus and the crista parotica of the petrosal, as well as between the incus and the malleus. Since the connection between these elements is lost later in post-hatching development, IF for beta-catenin was carried-out. Nuclear localised beta-catenin is a readout of canonical Wnt signalling, and is known to negatively regulate chondrocytes differentiation and promote joint formation. Few betacatenin positive cells were observed within the cartilage of the middle ear and petrosal at 0 days, though beta-catenin was strongly expressed in the neuro-epithelium of the inner ear. At post-hatching day 3, the incus and crista parotica were still fused, although the cells joining the two elements resembled fibrocartilage or immature chondrocytes. Expression of Sox9 was still strong and continuous throughout all elements, however Collagen Type 2 expression was weaker in the fusion region, possibly indicating a change in cartilage type from hyaline cartilage to fibrocartilage. Interestingly nuclear beta-catenin, suggestive of active Wnt signalling, was observed in two stripes, in the chondrocytes between the incus and petrosal, and within the malleus-incus joint, indicating suppression of cartilage fate in these regions. Upregulation of Wnt signalling between the incus and petrosal therefore, may play a role in formation of a joint between these two, initially fused, structures.

While the fusion between the incus and petrosal in Echidna and Platypus could be explained by the evolutionary distance between Monotremes and Therian Mammals, it has also been suggested that the incus is transiently attached to the cranial base in 7-week-old Human fetuses. This suggests that the potential for fusion may be a default state in Mammals. In order to examine this, Anthwal et al. next undertook fate mapping experiments in the Mouse, and investigated the relationship between the incus and petrosal in other Eutherian Mammals during embryonic development.
Sox9 expressing cells were fate mapped by tamoxifen induction at embryonic day 14.5 in Sox9CreERT2; tdTomato Mice, which were then collected at postnatal day 0. At this stage Sox9 (green) was expressed in the petrosal and incus and suspensory ligaments, overlapping with the red fluorescent protein marking the Sox9 lineage cells. In addition, the red Sox9 lineage cells were found in the Sox9 negative mesenchymal cells, in the gap between the petrosal and incus. A pre-cartilaginous bridge is therefore evident in the mouse between the incus and the crista parotica. Next, expression of Sox9 was investigated at embryonic day 14.5. The incus, and the crista parotica are both neural crest derived, while the rest of the petrosal is mesodermal. Anthwal et al. therefore looked at the expression of Sox9 (red) in Mesp1Cre;mTmG mice, where mesoderm-derived tissue can be detected by anti-gfp fl IF. Since tissue processing and wax embedding removes endogenous fluorescence, the membrane red fluorescent protein that is expressed in the non-mesodermal tissue of Mesp1Cre;mTmG mice was not detectable in these slides. Consequently, all red signal was Sox9 immunofluorescence staining. Sox9 protein was expressed continuously between the incus and the petrosal. The incus Sox9 expression domain was continuous with the expression domain of the neural crest -derived crista parotica, which in turn was fused to the mesodermal portion of the petrosal. Since the incus does not fuse with the petrosal in the mouse, despite the expression of Sox9 between the elements, we next looked at the mRNA expression of joint markers Gdf5 and Bapx1 between the incus and petrosal of mice by in situ hybridisation. Gdf5 was expressed in the mesenchyme between the incus and petrosal, as well as in the malleus-incus joint. Bapx1, which specifies both the malleus-incus joint and the quadrate-articular joint, was not expressed in between the incus and the petrosal. In the Mouse, therefore there is a potential for the incus and crista parotica to fuse but they are prevented from doing so by the upregulation of the joint marker Gdf5.

Very close associations between the incus and crista parotica during development were also observed in other Eutherian Mammals via PTA stained microCT, suggesting that interactions between these two elements are observed as a feature prenatally in Eutherian Mammals, similar to post-hatching monotremes. The function of this prenatal connection between the upper and lower jaw is unclear but may act as a brace to buffer movement during this period.
Next Anthwal et al. investigated the articulation between the incus and petrosal observed in the developing Opossum. It was originally suggested that the Marsupial incus forms a joint with the crista parotica, although this was disputed in Monodelphis. A previous study found no evidence of a joint but did show the mesenchyme between the crista parotica and incus as being condensed. Anthwal et al. therefore investigated the extra cellular matrix components of the mesenchyme surrounding the Opossum incus in more detail. It was noted that mesenchyme surrounding the crus breve and superior portion of the body of the incus had a more intense staining with alcian blue compared to those regions around the inferior border of the incus and the other ossicles. This pattern was observed throughout ossicle development. In order to further characterise the differences in the extra cellular matrix in the different regions of the middle ear mesenchyme, immunohistochemistry for versican was carried out. Versican is a large proteoglycan with side chains of glycosaminoglycans, such as hyaluronic acid. Proteoglycan complexes act to attract water, and are held in place by collagen fibres to stiffen the matrix in hyaline cartilage, and act to lubricate articular cartilage. Versican is required during the initial condensation of mesenchyme but is absent from mature cartilage, where aggrecan is expressed. Versican expression is maintained in the joint region during limb cartilage development, acting to inhibit maturation of the mesenchyme to form cartilage.

Versican was strongly expressed in the mesenchyme surrounding the short arm of the incus at 5 days, 10 day and 27 days, correlating with the region of strong alcian blue expression. The high level of versican around the crus breve therefore suggests a role for the extra cellular matrix in providing a buffering function in this region. Cell density of the mesenchyme was measured in regions with strong alcian blue/versican staining and compared against the cell density of regions with low alcian blue/versican staining. Unpaired two-tailed t-test demonstrated that the regions with high alcian blue had a significantly higher (p=0.0152) cell density than those regions with lower alcian staining.
Versican is processed by ADAMTS family members for clearing and remodelling. While the full-length form of versican is thought to have a structural role, the cleaved form has an active role in signalling, influencing morphogenesis and tissue remodelling. Interestingly when we analysed the cleaved form of versican, using antibodies against DPEAAE, the expression was largely reciprocal to that of uncleaved versican, with lower levels specifically around the crus breve. This suggests that versican around the incus is protected from cleavage allowing it to maintain its structural role. The lack of cleaved versican around the crus breve, suggests the lack of a signalling role in this region, in agreement with the low level of expression of CD44, a cell surface receptor and binding partner of versican-hyaluronan complexes. CD44 was not associated with the mesenchyme around the crus breve, but was instead restricted to the perichondrium of the cartilaginous elements and periosteum of the skeletal elements of the ear.
The incus of adult Mammals plays a key role in hearing. Anthwal et al.'s data here suggest that the incus also plays a transient role supporting the lower jaw against the cranial base during both Marsupial and Monotreme postnatal development. The incus and petrosal were found to be fused at hatching in both Monotremes. During this early fusion period, the puggle would be feeding exclusively on milk and Meckel’s cartilage could therefore act as a flexible elastic strut to facilitate jaw movement.

Interestingly, a potential role of the ear ossicles in jaw support was also observed in Eutherians during prenatal development. Fate mapping and gene expression studies in Mice indicated that the crus breve of the incus and the crista parotica were formed from a continuous region of Sox9 expressing chondrogenic cells, separated by expression of the joint marker Gdf5. Furthermore, the incus and cranial base temporarily fuse during the development of the Human middle ear region, and abut during bat development. Together these data indicate that the relationship of the incus to the cranial base is not a derived feature of Monotremes, and that the common Mammal-like Reptile ancestors of both Monotremes and Therian Mammals may have formed an articulation between the quadrate/incus and petrosal through fusion of the elements followed by joint formation though Wnt and Gdf5 signalling.
The current study indicates that the first pharyngeal arch-derived incus forms a continuous field of chondrocytes with the second arch-derived crista parotica, which in turn is fused with the mesoderm-derived body of the petrosal. The borders between these developmentally distinct populations are, therefore, not always reflected by the mature anatomy.
For young Monotremes and Marsupials, the middle ear must function as part of the mandible postnatally until the dentary-squamosal bones have formed. This is similar, but not identical to the situation in Cynodont ancestors of Mammals. In these animals, the quadrate/incus articulated with a number of cranial elements, including the quadratojugal, to stabilise the jaw articulation. These connections and many elements like the quadratojugal have been lost in extant Mammals in order to free the incus and increase its mobility during sound transmission. The mechanical requirements for feeding placed upon the middle ears in Monotremes and Marsupials during early life have resulted in the fusion of the incus and petrosal in monotremes, and the elongated contact supported by a proteoglycan matrix in Marsupials. These adaptations allow for stabilisation of the middle ear before the development of the dentary-squamosal joint and separation of the middle ear from the mandible, but do not compromise the effectiveness of the middle ear in later life.
The crus breve of the incus is elongated in the developing Opossum compared with other species analysed. In order to feed by suckling in the absence of a dentary-squamosal joint Anthwal et al. propose that this anatomy allows for an increased surface contact with the cranial base during postnatal development, which, in combination with the proteoglycan-rich surrounding mesenchyme, acts to stabilise the mandible against the rest of the head. It is noted that many adult Marsupials have a relatively elongated crus breve of the incus compared to Eutherian species, for example the Bare-tailed Woolly Opossum, Caluromys philander, and the Grey Short-tailed Opossum, Monodelphis domestica. Even when Eutherian Mammals have a longer crus breve, such as in Talpid Moles, the process is thinner and more finger-like compared to that of Marsupials. This may be a consequence of the developmental requirement for an elongated short process to facilitate feeding before the development of the mature Mammalian jaw articulation.
In the majority of adult Marsupials, including Monodelphis, the incus is suspended from the cranial base by suspensory ligaments, and the crus breve extends into a fossa. One interesting exception is the Marsupial Mole, in which the crus breve has a connective tissue attachment to a lamella on the petrosal. This results in the middle ear ossicles being affixed to the cranial base, an adaptation to a fossorial niche found in other Mammals such as in True Moles. In light of the current study, the absence of an incudal fossa in the Marsupial Mole may be interpreted as a retention of the juvenile petrosal morphology (paedomorphy).
In adult non-Mammalian Amniotes the homologue of the incus, the quadrate, and cranial base are strongly attached by fibrous syndesmoses or cartilaginous synchondroses, and Anthwal et al. show that a synovial joint appears to form in Geckos during development. In the neo-natal Opossum neither type of connection is observed. In neonatal Marsupials, the connection between the incus and petrosal has been described as being an ‘immature syndesmosis’, which acts as a ‘supportive strut’ during sucking. Anthwal et al. demonstrate a specialised condensed mesenchyme surrounds the incus of Opossum postnatal juveniles. They show that this condensed mesenchyme is rich in the proteoglycan versican. In contrast expression studies in Human foetuses demonstrate that versican is restricted to the perichondrium of Meckel’s cartilage, with high hyaluronic acid levels within the joints but not surrounding the incus. This concentration of versican around the crus breve therefore may be a feature of Monodelphis, and perhaps Marsupials in general.
The versican-rich mesenchyme may act to either stabilise the incus by increasing the tension of the surrounding mesenchyme during feeding, ‘lubricate’ the articulation between the incus and cranial base by increasing the hydration of the extra cellular matrix, or both. In keeping with this role, versican is dynamically expressed at the pubic symphysis during pregnancy in Mice, during which time the Mouse pubic symphysis forms a fibrous joint or syndesmosis. Significantly, there is little cleaved versican (DPEAAE) around the crus breve of the incus, suggesting a mechanical, rather than a signalling role. Overall it is likely that this mesenchyme is supporting the incus, rather than enabling mobilisation, with the high level of uncleaved versican acting to increase fibroviscocity while also elevating hydration of the extra cellular matrix. In this way, the mesenchyme around the incus acts as a cushion during the mechanical stress of suckling.
Meckel’s cartilage persists to at least 50 days post-hatching in the platypus. At this timepoint, juvenile Monotremes have two connections between the lower and upper jaw. The first connection is through the middle ear, which in juveniles remains attached to the mandible and articulates with the cranial base via the incus. The second is the later developing novel Mammalian jaw joint. Only much later in the life of the young does it appear that the connection between the middle ear and mandible is lost, and the malleus and incus act as a definitive Mammalian middle ear. The connection of the incus to the cranial base appears to be maintained in the adult Echidna and Platypus. This would be expected to impact on the movement of the incus, and therefore the efficiency of hearing, reflected in the poor hearing reported for Monotremes.
This novel finding of a double cranial articulation in the juvenile has significant implications for the evolution of the middle ear and jaw joint in Mammals. Fossil evidence indicates that Mammalian ancestors had a persistent connection between the middle ear ossicles and the jaw, as evidenced by the presence of an ossified Meckel’s element, or a dentary groove and post dentary trough, supporting a persistent Meckel’s cartilage. For these Animals, the connection of the middle ear with the jaw took one of two forms, in each case the Mammalian secondary jaw joint was present. The first was a more basal mandibular middle ear where the incus and malleus were firmly attached to the cranial base and dentary respectively. More derived fossils had a partial, or transitional Mammalian middle ear, where the middle ear was medially inflected away from the dentary, presumably allowing for improved vibration, but the malleus was still connected to the jaw, via Meckel’s cartilage. In these fossils with a partial Mammalian middle ear, little is understood of the rear of the ossicular chain, where the incus meets the petrosal, due to the poor and rare preservation of middle ear ossicles in the fossil record, a consequence of their small size. For example, only recently has a Multituberculate with a complete incus been described. Anthwal et al.'s data suggest that even in these transitional Mammals with a partial Mammalian middle ear, the incus would have still articulated with the cranial base via the crista parotica, at least at some point during the Animal’s life history.
The definitive Mammalian middle ear appears to have evolved independently in Monotremes and Therian Mammals. Due to the absence of evidence we do not know if the incus articulation in Animals with a partial Mammalian middle ear varied in a lineage specific manner, with the Therian lineage resembling juvenile Marsupials, and Monotremaformes resembling juvenile Platypuses and Echidna, or if both lineages had similar articulations. The data from transgenic reporter Mice, along with data from Humans suggests that the Monotreme-type fusion and articulation of the incus with the cranial base may have been common in Mammal like-Reptiles. As such, the developing Monotreme, with a double jaw articulation and a fused or articulated incus and petrosal, provides an exciting model for the study of the developmental basis of Mammalian evolution.
See also...





Online courses in Palaeontology.
Follow Sciency Thoughts on Facebook.
Follow Sciency Thoughts on Twitter.