
The term pathology refers to malformations of specimens of a population or species that are caused by exogenic or endogenic processes. Exogenic processes include injuries due to a predator attack, parasite infections, and the colonisation of the shell during lifetime. Endogenic processes include infections, mutations, or other illnesses. The term palaeopathology is used pathological features of fossils. To ensure accurate palaeoecological interpretations it is required to distinguish between pathologies and pseudopathologies, e.g. tectonically deformed Belemnite rostra. Fractures of tectonically deformed rostra are filled with blocky calcite cement instead of biogenic radial fibrous calcite. Such rostra, therefore, do not contribute to our understanding of Belemnite palaeoecology. The visible reactions of an affected specimen are called symptoms while several abnormalities that occur in a single specimen are described as complex syndromes. As a rule, only reactions to the disturbing factor should be considered to characterise pathological symptoms or syndromes. The disturbing factor (pathogen) underlying the palaeopathological features of fossil hardparts is only rarely identified. In exceptional cases, specific bite traces with a distinct shape or morphology of shell breakage are preserved and allow the identification of specific producers such as Crustaceans, Coleoids, Fishes, or Reptiles.
Pathologies of modern and fossil Cephalopods have received proportionally large attention. The first reports on malformed Cephalopods were made by Johann Samuel Schröter in 1774, although he did not recognise them as such. Later, pathological specimens were regarded as monstrous natural curiosities. Later, pathological specimens were regarded as monstrous natural curiosities. The potential factors causing abnormal morphologies were first discussed in the 1890s. Consequently, exploration of the disturbing factors was increasingly used to reconstruct the palaeobiology and palaeoecology of the affected specimens and helped in reconstructing phylogenetic processes.
Due to their internal shell, Coleoid Cephalopods such as Belemnites are not affected by epizoans that settle on the external shells of Nautiloids or Ammonoids. In externally shelled Cephalopods, damage to the phragmocone can result in uncontrolled flooding of the chambers, which in turn leads to lethal drowning by shell implosion. In contrast, the internal shell of Coleoids often remains in the mantle sac after being fractured. In case the animal survived the attack, damaged skeletal parts are often successfully repaired.
Data gained from palaeopathological studies of Cephalopods helped in estimating the efficiency of the buoyancy apparatus. Synecological factors can be reconstructed by the recognition of parasite infections or the presence of epizoa which in turn helped to reconstruct potential habitats and life habits. Lethal or sublethal injuries also add information on predator-prey interactions. Recent studies have provided a detailed overview on palaeopathological Belemnite rostra and introduced forma aegra-types for recurring kinds of abnormalities following the scheme developed for Ammonoids. It should be noted that forma aegra-types do not refer to taxonomic entities but are a type of open nomenclature for the classification of pathologies. For fossil coleoid cephalopods, including belemnites, three categories of palaeopathologies can be distinguished based on their causes: (i) regeneration of injuries including inflammatory infection often caused by an unsuccessful predatory attack, (ii) prod traces due to collisions, (iii) parasite infection.
Most malformations of Belemnite rostra resulted from mechanical injuries, often caused by failed predation attempts, and less commonly from parasite infestations. It has been argued that rostra broke due to mechanical stress induced during active digging in the sediment with the rostrum. Due to the likely nektic lifestyle of belemnites and the counterweight function of the rostrum that allows a horizontal swimming position, injuries due to digging are highly unlikely.
In a paper published in the journal Acta Palaeontologica Polonica on 24 March 2020, René Hoffmann and Kevin Stevens of the Institut für Geologie, Mineralogie und Geophysik at the Ruhr-Universität Bochum, Marie-Claire Picollier of St. Pantaly d’Excideuil, Jörg Mutterlose, also of the Institut für Geologie, Mineralogie und Geophysik at the Ruhr-Universität Bochum, and Christian Klug of the Paläontologisches Institut und Museum at Universität Zürich, demonstrate that nano-computed-tomography scanning of pathological Belemnite rostra provide data with an excellent resolution, allowing a detailed description of internal features including malformations.
High resolution and good signal to noise ratios in the resulting computed-tomography-images are important for the successful study of pathological belemnite rostra because of the continuous deposition of concentric growth increments. That mode of growth resulted in a successive attenuation of the primary morphology of the injury and the resulting symptom due to the deposition of post-traumatic growth increments. Based on computed-tomography-data, Hoffmann et al. provide short descriptions of the different types of palaeopathologies and discuss their potential causes. Furthermore, they demonstrate additional applications such as the recognition of a heterogeneous or homogenous composition of Belemnite rostra due to diagenesis, sedimentary fill, structures of the rostrum surface, and internal structures of the phragmocone such as septal spacing and the position of the siphuncle.
A total of 18 pathological belemnite rostra from Lower Jurassic (Toarcian) to Upper Cretaceous (Campanian) deposits, were computed-tomography-scanned. The material can be attributed to the following genera (in alphabetical order, number of studied specimens in parenthesis): ?Acrocoelites sp. (1), Belemnellocamax spp. (4), Belemnitella sp. (2), Duvalia emerici(1), Goniocamax sp. (1), Gonioteuthis spp. (5), Hibolithes jaculoides (2), Neoclavibelus subclavatus (1), and Pseudobelus sp. (1).
Hoffmann et al. describe various pathologies encountered according to their morphological expressions including two-tipped specimens, blisters and pearls, broken rostra, which healed straight, rostra with a kink or bent rostra. Differences in grey scale values in the images suggest different materials such as calcite, silica, pyrite, or sediment. The growth increments of the rostrum, for example, are visible in computed-tomography-images due to the varying amount of organic matter, which has significant lower absorption properties compared to pure calcite. This implies that organic-rich growth increments are darker compared to those composed of pure calcite.

One ?Acrocoelites sp. specimen (PIMUZ 37346) and one Gonioteuthis sp. specimen (RUB-Pal 11264) rostrum are characterised by two divergent apices. In both specimens one apex represents the continuation of the original rostrum axis and bears radial striation on its outer surface. The second apex of ?Acrocoelites sp. bears one strong dorsal furrow. Superficially, the arrangement of the ?Acrocoelites sp. apices resembles a crustacean claw. Unfortunately, the computed-tomography-data did not reveal the course of growth increments or other internal structures, which suggests an irregular formation of the skeletal calcite.

The continuation of the normal rostrum growth in the Gonioteuthis sp. is supported by the presence of the apical line and growth increments. The rostrum surface of this Gonioteuthis sp. partially shows dissolution features or poorly mineralized layers exposing triangular structures. Some of these features suggest the presence of meandering trace fossils. Both apices were partially hollow and subsequently filled with sediment. Development of the two apices is reflected in the image series based on virtual cross sections. Areas darker than the calcitic parts of the rostrum and mostly restricted to the rostrum surface indicate the presence of silica.
One Neoclavibelus subclavatus (SNSB-BSPG-83264) and one Belemnitella sp. (RE 551.763.333. A 5238) rostrum each show a bump-like structure. Virtual cross sections of the Neoclavibelus subclavatus rostrum reveal the presence of sediment inside the rostrum. This debris was subsequently overgrown by calcite precipitated by the Belemnite, causing the malformation. The longitudinal section shows that the contaminant covers parts of the juvenile rostrum, thereby causing a deviation in growth increment orientation. White areas suggest the presence of pyrite, which completely absorbs the X-rays. The bump-like structure in the Belemnitella sp. rostrum was probably caused by a similar process. In this case, the foreign material caused the bending of dorsolateral depressions covering the rostrum surface. Darker areas along the margin of the rostrum suggest the presence of silica. For both species, Hoffmann et al. also provide high-resolution computed-tomography-images. A large part of the Goniocamax sp. rostrum shows signs of exfoliation. Hoffmann et al.'s computed-tomography-based examination of the area where outer rostral layers thin out reveals, that sediment was deposited between distinct growth increments of the belemnite rostrum. Both specimens show hollow areas, partially filled with carbonate crystals, within the rostrum forming blisters. The Goniocamax sp. blister is elongated with a spiral morphology. The sediment filled blister contrasts with the massive bump-like structures of another two specimens, undetermined specimen CASP K9068 and Duvalia emerici. The undetermined rostrum shows a distinctive layer from which the growth perturbation starts in longitudinal- and cross section. This structure resembles the phenomenon described for Neoclavibelus subclavatus and Belemnitella sp. For Duvalia emerici, no such perturbation has been identified.

One Belemnite assigned to Gonioteuthis sp. (RUB-Pal 11301) and one assigned to Hibolithes sp. (RUB-Pal 11303) have fractured rostra. Due to strong silification, the results from the computed-tomography-scans remain inconclusive for the Gonioteuthis sp. rostrum. The apical line appears to form a continuous line without the formation of a blunt rostrum. Parts of the rostrum increments seem to be poorly mineralised, causing parts of the juvenile rostrum to stick out where normally the alveolus is situated. This example demonstrates the potential of the computed-tomography technique to better understand the mineralogical composition of Belemnite rostra. In Hoffmann et al.'s specimen silification rings correspond to darker areas in virtual section images. Virtual sections of the Hibolithes jaculoides rostrum reveal a broken juvenile rostrum with the apex missing. The fracture was overgrown by irregularly formed rostrum material. While the outer layers show regularly formed growth increments, the internal part appears to be chaotic without forming an apical line. This irregular growth results in a rostrum morphology comparable to the genus Produvalia.

Two specimens of Gonioteuthis spp. display a marked distortion of the rostrum. Both, the rostrum and the phragmocone were affected in one specimen, post mortem overgrown by an Oyster In this case parts of the juvenile rostrum are not preserved (RUB-Pal 11302). The rostrum surface shows faint radial striae. The second specimen (SNSB-BSPG-83246) yields an intact phragmocone but parts of the juvenile rostrum broke off. The phragmocone is partially filled with pyrite (white area) and sediment (irregular greyish). Due to partial silification, the growth increments are well visible. Areas with strong bending are silicified, and the rostrum surface is rough due to dissolution, poor mineralisation, or both. Although strongly bent, the rostrum shows growth increments and an apical line.

One rostrum each of Gonioteuthis sp. and Pseudobelus sp. are strongly distorted resulting in a change of growth direction. The specimen of Gonioteuthis sp. (SNSBBSPG-83370) deviates from the normal (straight) growth direction by about 90° with its apex pointing slightly towards the alveolus. A small part of the phragmocone, now filled with sediment, is preserved and the rostrum is heavily silicified. A few growth increments and a faint trace of the apical line are visible. The rostrum of Pseudobelus sp. (RUB-Pal 3196) shows the strongest deformation reported here. Computed-tomography-data suggest three traumatic events resulting in fractures and finally a reversion of the growth direction towards the head of the animal. Growth increments and apical line are, however, well visible in the longitudinal section. The cross-section image shows four growth zones representing a temporal sequence implying that the rostrum broke during four successive and independent events.

Four malformed specimens of Belemnellocamax sp. with fractured rostra show weak to strong reactions that occurred during the healing process. One specimen (GM 04.1918) is a knee-like bent rostrum with a spongy, highly porous internal structure. A second specimen (GM 02.1918) has a short rostrum with a blunt apex due to a spherical depression. The virtual section reveals a broken juvenile rostrum associated with a porous area surrounding it. The third specimen (GM 01.1918) also has a broken juvenile septum, which extended beyond growth increments of older parts of the rostrum. Subsequently formed growth increments never covered the juvenile rostrum entirely, forming a callus-like structure. Parts of the alveolar region are preserved allowing for a correct orientation of the rostrum. Specimen four (GM 03.1918) shows a slightly irregular rostrum morphology with curved depressions and a small hole on the rostrum surface. Virtual longitudinal sections reveal that the hole at the surface is connected to a canal that appears to start in the central alveolar region. Growth increments, otherwise well visible, disappear around the canal.

Like all Coleoid Cephalopods, Belemnites formed their internal shell within the shell sac. Belemnite rostra were secreted by the muscular mantle that is fused above the shell mantle in Endocochleate Cephalopods. Growth increments are rhythmically added to the rostrum by simultaneous accretion on its surface. This mode of biomineralisation is comparable to the formation of the cuttlebone sheath of Sepiids including the spine, which contains high amounts of organic material. The rostrum is composed of radiaxial calcite fibres that cross the concentric growth increments.

The recent application of non-invasive imaging methods other than magnetic resonance imaging to Belemnite rostra, such as synchrotron radiation based micro-computed-tomography, revealed the presence of a new rostral ultrastructure, suggesting a composition of two distinct calcite phases. Micro-computed-tomography has previously been used to describe bioerosion features (trace fossils), which are often found on Belemnite rostra, and to determine rostrum volumes to reconstruct temporal changes in Belemnite size. The description of a variety of pathological phenomena highlights the advantage of the higher spatial resolution and suitable contrast properties accessible by computed-tomography-derived volume data to explore the three-dimensional nature of pathological specimens.

The following malformation categories were used to group the various types of malformation:
Apex malformation describes a duplication or multiplication (up to five apices known so far) of the rostrum apex. It has been argued that due to traumatic events (mechani cal injuries) or the activity of parasites parts of the apex forming mantle epithelium were separated into locally independent secretion centres that formed several apices. Hoffmann et al. report apex malformations in one specimen of ?Acrocoelites sp. and Gonioteuthis sp. each.
Blister malformation includes blister thickenings of the rostrum often enclosing a hollow space, which may indicate the earlier presence of endoparasites. The locally proliferating mineralization of the rostrum can in rare cases lead to the formation of a second rostrum paralleling the primary rostrum. Hoffmann et al. report blister malformations in one specimen of Neoclavibelus subclavatus, Belemnitella sp., Hibolithes jaculoides, Goniocamax sp., an undetermined Belemnite, and Duvalia emerici each.
Blunt rostra malformation contains more or less club-shaped rostra that are thicker and shorter compared to normal specimens. They are the result of broken juvenile rostra. Depending on the timing and severity of the fracture, slightly to heavily deformed rostra result. Often, the earliest parts of the phragmocone including the initial chamber are affected, Extreme shortenings of the rostrum results in a droplet-shaped morphology. Hoffmann et al. report blunt rostra malformation in one specimen of Gonioteuthis sp., one specimen of Hibolithes jaculoides, and two Belemnellocamax sp. specimens.
Bent rostra malformation is characterised by multiple rostrum fractures. The dislocated rostrum fragments were covered by post-traumatic growth increments. Depending on the size and position of the fragments within the mantle sac, distortions of variable angles (within a plane or a screw-like arrangement of the fragments in different planes) resulted. This phenomenon includes specimens with fragments grown together sub-parallel to each other after the attack. Hoffmann et al. report bent rostra malformation in two specimens of Gonioteuthis spp. and one specimen of Belemnellocamax sp.
Hook-shaped rostra are often the result of rostrum fractures that occurred during early ontogeny. Parts of the broken rostrum become dislocated and post-traumatic growth is preferentially oriented towards one side of the rostrum resulting in a change of growth direction. In cases where the rostrum becomes significantly shorter, a form transitional between the hook-shaped rostra and collar formation exists. A 180° reversal of the growth direction of the rostrum towards the head of the animal due to several fractures is documented by Hoffmann et al. for the first time. Hoffmann et al. report Hoffmann et al. in one specimen of Gonioteuthis sp. and Pseudobelus sp. each.
Sceptre-shaped rostra occur where a mechanical injury of the mantle sac epithelium leads to a partial lack of growth increments. This is related to the disabled function of the injured epithelium to secrete shell material. This symptom is often observed in the apical area of the rostrum causing an incomplete apex. In this case juvenile, pre-traumatic parts of the rostrum form the apex. A damage of the lateral epithelium is related to a thinning of the rostrum in the affected area and result in a sceptre-shaped rostrum. Hoffmann et al. report sceptre-shaped rostra in one specimen of Gonioteuthis sp.
Collar formation deformation describes flap-like outgrowths at the sub-apical rostrum area without visible injury. This symptom has been reported from a variety of Jurassic and Cretaceous Belemnites and possibly results from a parasitic infestation This infestation caused the formation of a collar that is regarded as an effluence channel and potentially indicates an inflammation of the mantle sac epithelium. Hoffmann et al. report collar formation in one specimen of Belemnellocamax sp.
Most studies about modern Molluscan parasites deal with parasites of Bivalves. Bivalves host a diverse group of parasites and disease-causing agents such as, e.g., Viruses, Prokaryotes, Fungi, Protists, Parazoans, and Metazoans (Platyhelminths, Annelids, Molluscs, Bryozoans, and Arthropods). The most abundant parasites are endoparasitic Flukes (Trematoda, Digenea), which have also been observed in Gastropods. Larvae of Digenean Trematodes (Gymnophallidae) sometimes lead to the formation of blisters and have been found in fossil Bivalves dating back to the Triassic. Blister formation, however, can have a variety of causes, some of which are not due to parasites. Identification of a specific parasite taxon, however, is hampered by the fact that parasites are typically small-bodied (submillimetre) and usually lack biomineralised hardparts.
In modern Cephalopods, parasites are known from many species from all major oceans ranging from coastal, shelf, to oceanic and deep-sea environments. This is no surprise given that Cephalopods are a key trophic element in marine ecosystems. One study documented 200 species of endoparasites in Cephalopods with macroparasites such as Nematodes, Copepods, and Isopods. These predominantly affect non-mineralized tissues such as the digestive tract and the gills. The infestation of mainly non-mineralised tissues limits the fossil preservation potential of parasite activity and the resulting host reaction.
For Belemnites, most reported pathologies are due to survived predator attacks (e.g. bent rostra), a few malformations are assigned to the activity of parasites like, blister malformation, collar formation, and apex malformation. Although Hoffmann et al. identify a parasite infection as the most likely trigger of these pathologies, attributing the deformations to a specific parasite taxon is difficult to impossible. While some parasites leave characteristic patterns on their host, this is not necessarily definitive evidence of their presence. Different parasites can leave similar traces and taxonomically distant parasites can inflict similar symptoms on their hosts because of convergence in the evolution of host-exploitation strategies. Differential diagnosis for palaeopathological phenomena may lead to the conclusion that two or more alternative conditions remain as potential triggers.
Hoffmann et al.'s high-resolution computed-tomography-data represent an innovative and non-destructive way to analyse the internal structure of Belemnite rostra. computed-tomography-images revealed internal structures that allow the examination of a variety of pathological phenomena and to differentiate between different types of pathology. Most importantly, the computed-tomography-data allow to infer potential causes of these deformations (predator attack or parasite activity), therefore going beyond a classification of pathologies from the surface of a specimen alone. Hoffmann et al. thereby demonstrated the usefulness and feasibility of the application of non-invasive imaging methods to the field of palaeopathology. The data gained can be further used for the identification of various diagenetic alteration features in Belemnite rostra. computed-tomography-data turned out to perform better than magnetic resonance imaging-data in terms of spatial resolution and contrast.
See also...
A total of 18 pathological belemnite rostra from Lower Jurassic (Toarcian) to Upper Cretaceous (Campanian) deposits, were computed-tomography-scanned. The material can be attributed to the following genera (in alphabetical order, number of studied specimens in parenthesis): ?Acrocoelites sp. (1), Belemnellocamax spp. (4), Belemnitella sp. (2), Duvalia emerici(1), Goniocamax sp. (1), Gonioteuthis spp. (5), Hibolithes jaculoides (2), Neoclavibelus subclavatus (1), and Pseudobelus sp. (1).
Hoffmann et al. describe various pathologies encountered according to their morphological expressions including two-tipped specimens, blisters and pearls, broken rostra, which healed straight, rostra with a kink or bent rostra. Differences in grey scale values in the images suggest different materials such as calcite, silica, pyrite, or sediment. The growth increments of the rostrum, for example, are visible in computed-tomography-images due to the varying amount of organic matter, which has significant lower absorption properties compared to pure calcite. This implies that organic-rich growth increments are darker compared to those composed of pure calcite.

Rostrum of Belemnite ?Acrocoelites sp., PIMUZ 37346, Toarcian, Altdorf (southwest-Germany) with apex malformation. (A) Overview (A₁), (A₂). (B) Close-up of the two apices, showing the radial furrows covering the apex that represent the 'normal' tip of the rostrum (B₁)–(B₅). Hoffmann et al. (2020).
One ?Acrocoelites sp. specimen (PIMUZ 37346) and one Gonioteuthis sp. specimen (RUB-Pal 11264) rostrum are characterised by two divergent apices. In both specimens one apex represents the continuation of the original rostrum axis and bears radial striation on its outer surface. The second apex of ?Acrocoelites sp. bears one strong dorsal furrow. Superficially, the arrangement of the ?Acrocoelites sp. apices resembles a crustacean claw. Unfortunately, the computed-tomography-data did not reveal the course of growth increments or other internal structures, which suggests an irregular formation of the skeletal calcite.
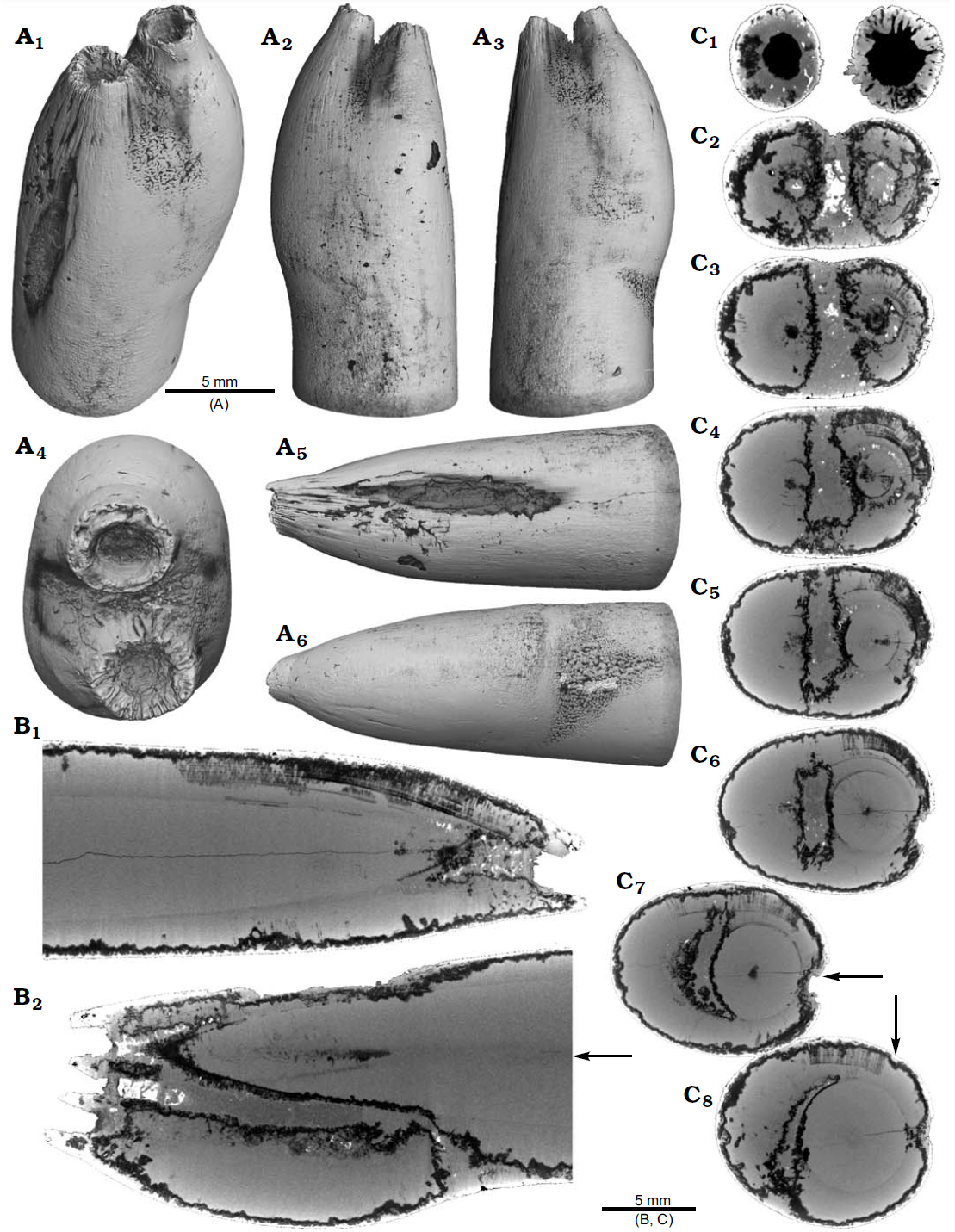
Rostrum of Belemnite Gonioteuthis sp., RUB-Pal 11264, Campanian, Höver (northwest-Germany) with apex malformation. (A) Surface images showing the hollow opening of the two apices, and the partially dissolved or poorly mineralized rostrum (A₁)–(A₆); arrow in (A₅) indicates meandering trace fossil. (B) Longitudinal section showing growth increments, apical line, and irregular mineralized layers (B₁), (B₂). (C) Cross sections with increasing distance to the apices (C₁)–(C₈), showing the process of apex separation, only the straight continuous apex (arrows) develops distinct growth increments and the apical line (see also the arrow in B₂), note: the area between the apices is filled with sediment (diffuse grey area) and pyrite (white area), black area silicified. Hoffmann et al. (2020).
The continuation of the normal rostrum growth in the Gonioteuthis sp. is supported by the presence of the apical line and growth increments. The rostrum surface of this Gonioteuthis sp. partially shows dissolution features or poorly mineralized layers exposing triangular structures. Some of these features suggest the presence of meandering trace fossils. Both apices were partially hollow and subsequently filled with sediment. Development of the two apices is reflected in the image series based on virtual cross sections. Areas darker than the calcitic parts of the rostrum and mostly restricted to the rostrum surface indicate the presence of silica.
One Neoclavibelus subclavatus (SNSB-BSPG-83264) and one Belemnitella sp. (RE 551.763.333. A 5238) rostrum each show a bump-like structure. Virtual cross sections of the Neoclavibelus subclavatus rostrum reveal the presence of sediment inside the rostrum. This debris was subsequently overgrown by calcite precipitated by the Belemnite, causing the malformation. The longitudinal section shows that the contaminant covers parts of the juvenile rostrum, thereby causing a deviation in growth increment orientation. White areas suggest the presence of pyrite, which completely absorbs the X-rays. The bump-like structure in the Belemnitella sp. rostrum was probably caused by a similar process. In this case, the foreign material caused the bending of dorsolateral depressions covering the rostrum surface. Darker areas along the margin of the rostrum suggest the presence of silica. For both species, Hoffmann et al. also provide high-resolution computed-tomography-images. A large part of the Goniocamax sp. rostrum shows signs of exfoliation. Hoffmann et al.'s computed-tomography-based examination of the area where outer rostral layers thin out reveals, that sediment was deposited between distinct growth increments of the belemnite rostrum. Both specimens show hollow areas, partially filled with carbonate crystals, within the rostrum forming blisters. The Goniocamax sp. blister is elongated with a spiral morphology. The sediment filled blister contrasts with the massive bump-like structures of another two specimens, undetermined specimen CASP K9068 and Duvalia emerici. The undetermined rostrum shows a distinctive layer from which the growth perturbation starts in longitudinal- and cross section. This structure resembles the phenomenon described for Neoclavibelus subclavatus and Belemnitella sp. For Duvalia emerici, no such perturbation has been identified.

(A) Rostrum of Belemnite Neoclavibelus subclavatus, SNSB-BSPG-83264, Toarcian, Mistelgau (southwest-Germany) with blister malformation. Surface images showing the bump-shaped irregular rostrum growth (A₁)–(A₄); longitudinal sections showing presence of sediment (diffuse grey) and pyrite (white) within the rostrum, and increasing irregular growth increments (A₅), (A₆). (B) Rostrum of Belemnite Belemnitella sp., RE 551.763.333 A 5238, Late Cretaceous, northwest-Germany with forma aegra bullata (coll. Baschin). Surface images showing the bump-shaped irregular rostrum growth and imprints of an organic network on the rostrum surface in dorsolateral (B₁), (B₂) and ventrolateral (B₃), (B₄) views; longitudinal sections (B₅), (B₆), and cross section (B₇). (B₅)–(B₇) showing silification (black) along the rostrum surface but also along the malformed area. Hoffmann et al. (2020).
One Belemnite assigned to Gonioteuthis sp. (RUB-Pal 11301) and one assigned to Hibolithes sp. (RUB-Pal 11303) have fractured rostra. Due to strong silification, the results from the computed-tomography-scans remain inconclusive for the Gonioteuthis sp. rostrum. The apical line appears to form a continuous line without the formation of a blunt rostrum. Parts of the rostrum increments seem to be poorly mineralised, causing parts of the juvenile rostrum to stick out where normally the alveolus is situated. This example demonstrates the potential of the computed-tomography technique to better understand the mineralogical composition of Belemnite rostra. In Hoffmann et al.'s specimen silification rings correspond to darker areas in virtual section images. Virtual sections of the Hibolithes jaculoides rostrum reveal a broken juvenile rostrum with the apex missing. The fracture was overgrown by irregularly formed rostrum material. While the outer layers show regularly formed growth increments, the internal part appears to be chaotic without forming an apical line. This irregular growth results in a rostrum morphology comparable to the genus Produvalia.

(A) Rostrum of Belemnite cf. Hibolithes jaculoides, SNSB-BSPG-83251, Hauterivian, Heligoland (north-Germany) with blister malformation. Surface images showing the bump-shaped irregular rostrum growth, in lateral (A₁), dorsal (A₂), lateral (A₃), and ventral (A₄) views. Longitudinal section showing open pore space partially filled with carbonate cements (A₅). (B) Rostrum of Belemnite Goniocamax sp., MGUH 32024, Santonian, Bavnodde (Denmark) with blister malformation. Surface images of the rostrum showing the bump-shaped irregular rostrum growth (B₁)–(B₄). Longitudinal section with growth increments, apical line, and the bump-like structure with pore space (black) (B₅). Cross section through the malformed area showing the pore space partially filled with carbonate cement (B₆). Marginal section through the malformed area (B₇). (C) Undetermined Belemnite, CASP K9068, Greenland, Early Cretaceous with blister malformation. Surface images of the rostrum showing the irregular rostrum area (C₁)–(C₃). Longitudinal section showing growth increments, apical line and a distinct growth increment at which the malformation starts (C₄). Cross section that shows the distinct growth increment, and a horizontal fracture (white), as well as grains of pyrite (white) (C₅). (D) Rostrum of Belemnite Duvalia emerici, RUB-Pal 22170, Late Valanginian, Laborel (France) with blister malformation. Surface images of the rostrum showing the irregular rostrum area (half-pearl shaped), in ventral (D₁), lateral (D₂), (D₃), and dorsal (D₄) views. Longitudinal section through the malformed area (D₅). Cross section, both section reveal that the structure is massive, i.e., not a blister pearl (D₆). Hoffmann et al. (2020).
Two specimens of Gonioteuthis spp. display a marked distortion of the rostrum. Both, the rostrum and the phragmocone were affected in one specimen, post mortem overgrown by an Oyster In this case parts of the juvenile rostrum are not preserved (RUB-Pal 11302). The rostrum surface shows faint radial striae. The second specimen (SNSB-BSPG-83246) yields an intact phragmocone but parts of the juvenile rostrum broke off. The phragmocone is partially filled with pyrite (white area) and sediment (irregular greyish). Due to partial silification, the growth increments are well visible. Areas with strong bending are silicified, and the rostrum surface is rough due to dissolution, poor mineralisation, or both. Although strongly bent, the rostrum shows growth increments and an apical line.

A. Rostrum of Belemnite Gonioteuthis sp., RUB-Pal 11301, Campanian, Höver (northwest-Germany) with blunt rostra. Surface images showing no irregularities except for silification rings (A₁)–(A₃). Longitudinal section showing a homogenous internal rostrum with a dark layer along its outer margin (silica) (A₄). Volume rendering image shows a darker centre due to the maximum thickness of the structure but no additional features (A₅). Detail of the rostrum surface showing silification rings (A₆). Cross section with a homogeneous centre and a dark margin (A₇). (B) Rostrum of Belemnite Hibolithes jaculoides, RUB-Pal 11303, Hauterivian, Resse (northwest-Germany) with blunt rostra. Surface images showing the overall irregular rostrum morphology and the blunt and hollow apex in lateral (B₁), (B₂), ventral (B₃), and dorsal (B₄), apical (B₇) views. Longitudinal sections (central, subcentral) showing the broken juvenile rostrum, parts of the preserved phragmocone, and notable the lack of the apical line after the injury took place, note the irregular outline of the hollow central canal (B₅), (B₆). Cross section with the juvenile rostrum, and subsequently deposited homogeneous material, white areas indicate the presence of pyrite (B₈). Hoffmann et al. (2020).
One rostrum each of Gonioteuthis sp. and Pseudobelus sp. are strongly distorted resulting in a change of growth direction. The specimen of Gonioteuthis sp. (SNSBBSPG-83370) deviates from the normal (straight) growth direction by about 90° with its apex pointing slightly towards the alveolus. A small part of the phragmocone, now filled with sediment, is preserved and the rostrum is heavily silicified. A few growth increments and a faint trace of the apical line are visible. The rostrum of Pseudobelus sp. (RUB-Pal 3196) shows the strongest deformation reported here. Computed-tomography-data suggest three traumatic events resulting in fractures and finally a reversion of the growth direction towards the head of the animal. Growth increments and apical line are, however, well visible in the longitudinal section. The cross-section image shows four growth zones representing a temporal sequence implying that the rostrum broke during four successive and independent events.

Rostra of Belemnite Gonioteuthis spp. (A) RUB-Pal 11302, Campanian, Höver (northwest-Germany) with bent rostra. Surface images showing the knee-like morphology of the rostrum, and the attachment-base of an Oyster, in lateral (A₁), (A₄), dorsal (A₂), and ventral (A₄) views; note the weak furrows in (A₃). Median sections perpendicular to each other showing silicified areas (darker) and the broken phragmocone now filled with sediment, no additional internal feature visible (A₅), (A₆). (B) SNSB-BSPG-83246, Campanian, Höver (northwest-Germany) with bent rostra. Surface images showing the knee-like morphology of the rostrum, in ventral (B₁), lateral (B₂), (B₄), and dorsal (B₅) views. Cross section (B₃). Median sections showing silicified areas specifically at places heavily bent (darker) (B₆), (B₇); see also (A₅), (A₆) for the same phenomenon. Black box indicates close up in (B₇), showing the broken juvenile rostrum with growth increment (bent rostra), and the phragmocone partially filled with pyrite (white). Hoffmann et al. (2020).
Four malformed specimens of Belemnellocamax sp. with fractured rostra show weak to strong reactions that occurred during the healing process. One specimen (GM 04.1918) is a knee-like bent rostrum with a spongy, highly porous internal structure. A second specimen (GM 02.1918) has a short rostrum with a blunt apex due to a spherical depression. The virtual section reveals a broken juvenile rostrum associated with a porous area surrounding it. The third specimen (GM 01.1918) also has a broken juvenile septum, which extended beyond growth increments of older parts of the rostrum. Subsequently formed growth increments never covered the juvenile rostrum entirely, forming a callus-like structure. Parts of the alveolar region are preserved allowing for a correct orientation of the rostrum. Specimen four (GM 03.1918) shows a slightly irregular rostrum morphology with curved depressions and a small hole on the rostrum surface. Virtual longitudinal sections reveal that the hole at the surface is connected to a canal that appears to start in the central alveolar region. Growth increments, otherwise well visible, disappear around the canal.

(A) Rostrum of Belemnite Gonioteuthis sp., SNSB-BSPG-83370, Campanian, Höver (northwest-Germany) with hook-shaped rostra. Surface images showing the knee-like strongly bent and irregular morphology of the rostrum (A₁)–(A₄). Median section showing the irregular internal silification of the rostrum indicating poorly mineralized areas, growth and apical line partially visible (A₅). Volume rendering image with the darkest areas represented by the thickest or densest areas (A₆). (B) Rostrum of Belemnite Pseudobelus sp., RUB-Pal 3196, Valanginian, Barret-Meouge (France) with hook-shaped rostra. Surface images showing the strongly bent and irregular morphology of the rostrum with the apex growth in anterior direction (B₃)–(B₆). Median section overview and close up showing growth increments and the presence of pyrite along the apical line (white) (B₁), (B₇), (B₈). Cross section showing four growth center representing a temporal sequence (B₂). Hoffmann et al. (2020).
Like all Coleoid Cephalopods, Belemnites formed their internal shell within the shell sac. Belemnite rostra were secreted by the muscular mantle that is fused above the shell mantle in Endocochleate Cephalopods. Growth increments are rhythmically added to the rostrum by simultaneous accretion on its surface. This mode of biomineralisation is comparable to the formation of the cuttlebone sheath of Sepiids including the spine, which contains high amounts of organic material. The rostrum is composed of radiaxial calcite fibres that cross the concentric growth increments.

Rostra of Belemnite Belemnellocamax spp., Campanian, Ivo Klack (Sweden). (A) GM 04.1918, collar formation. Surface images showing the knee-like morphology of the rostrum (A₁)–(A₄). Longitudinal section showing the porous apical area (black) (A₅). Alveolar region (A₆). Cross section showing pore space within the rostrum (black) (A₇). (B) GM 02.1918, collar formation. Alveolar region (B₁). Cross section with the juvenile rostrum in its centre (B₂). Rostrum surface of the short, stout rostrum with a blunt apex and a concave impression (?dissolution feature), in lateral (B₃)–(B₆) and apical (B₇) views. Central and subcentral longitudinal section with the juvenile rostrum and the surrounding porous area (B₈), (B₉). Hoffmann et al. (2020).
The recent application of non-invasive imaging methods other than magnetic resonance imaging to Belemnite rostra, such as synchrotron radiation based micro-computed-tomography, revealed the presence of a new rostral ultrastructure, suggesting a composition of two distinct calcite phases. Micro-computed-tomography has previously been used to describe bioerosion features (trace fossils), which are often found on Belemnite rostra, and to determine rostrum volumes to reconstruct temporal changes in Belemnite size. The description of a variety of pathological phenomena highlights the advantage of the higher spatial resolution and suitable contrast properties accessible by computed-tomography-derived volume data to explore the three-dimensional nature of pathological specimens.

Rostra of belemnite Belemnellocamax spp. (A) GM 01.1918, Campanian, Ivo Klack (Sweden) with collar formation. Surface images showing the collar-like structure and parts of the juvenile rostrum (A₁)–(A₄). Longitudinal section (A₅) showing the sediment filled phragmocone with the protoconch, the Klähn’sche plane, the juvenile rostrum, and the material added after the traumatic event forming a collar-like structure but not covering the juvenile rostrum completely. Alveolar region (A₆). Opposite side with the juvenile rostrum sticking out (A₇). (B) GM 03.1918, Campanian, Ugnsmunnarna (Sweden) with fcollar formation. Irregular outline morphology of the rostrum (B₁)–(B₄). Longitudinal section to show the canal, growth increments (B₅), (B₆). Cross section through the apical line and the canal (B₇). Hoffmann et al. (2020).
The following malformation categories were used to group the various types of malformation:
Apex malformation describes a duplication or multiplication (up to five apices known so far) of the rostrum apex. It has been argued that due to traumatic events (mechani cal injuries) or the activity of parasites parts of the apex forming mantle epithelium were separated into locally independent secretion centres that formed several apices. Hoffmann et al. report apex malformations in one specimen of ?Acrocoelites sp. and Gonioteuthis sp. each.
Blister malformation includes blister thickenings of the rostrum often enclosing a hollow space, which may indicate the earlier presence of endoparasites. The locally proliferating mineralization of the rostrum can in rare cases lead to the formation of a second rostrum paralleling the primary rostrum. Hoffmann et al. report blister malformations in one specimen of Neoclavibelus subclavatus, Belemnitella sp., Hibolithes jaculoides, Goniocamax sp., an undetermined Belemnite, and Duvalia emerici each.
Blunt rostra malformation contains more or less club-shaped rostra that are thicker and shorter compared to normal specimens. They are the result of broken juvenile rostra. Depending on the timing and severity of the fracture, slightly to heavily deformed rostra result. Often, the earliest parts of the phragmocone including the initial chamber are affected, Extreme shortenings of the rostrum results in a droplet-shaped morphology. Hoffmann et al. report blunt rostra malformation in one specimen of Gonioteuthis sp., one specimen of Hibolithes jaculoides, and two Belemnellocamax sp. specimens.
Bent rostra malformation is characterised by multiple rostrum fractures. The dislocated rostrum fragments were covered by post-traumatic growth increments. Depending on the size and position of the fragments within the mantle sac, distortions of variable angles (within a plane or a screw-like arrangement of the fragments in different planes) resulted. This phenomenon includes specimens with fragments grown together sub-parallel to each other after the attack. Hoffmann et al. report bent rostra malformation in two specimens of Gonioteuthis spp. and one specimen of Belemnellocamax sp.
Hook-shaped rostra are often the result of rostrum fractures that occurred during early ontogeny. Parts of the broken rostrum become dislocated and post-traumatic growth is preferentially oriented towards one side of the rostrum resulting in a change of growth direction. In cases where the rostrum becomes significantly shorter, a form transitional between the hook-shaped rostra and collar formation exists. A 180° reversal of the growth direction of the rostrum towards the head of the animal due to several fractures is documented by Hoffmann et al. for the first time. Hoffmann et al. report Hoffmann et al. in one specimen of Gonioteuthis sp. and Pseudobelus sp. each.
Sceptre-shaped rostra occur where a mechanical injury of the mantle sac epithelium leads to a partial lack of growth increments. This is related to the disabled function of the injured epithelium to secrete shell material. This symptom is often observed in the apical area of the rostrum causing an incomplete apex. In this case juvenile, pre-traumatic parts of the rostrum form the apex. A damage of the lateral epithelium is related to a thinning of the rostrum in the affected area and result in a sceptre-shaped rostrum. Hoffmann et al. report sceptre-shaped rostra in one specimen of Gonioteuthis sp.
Collar formation deformation describes flap-like outgrowths at the sub-apical rostrum area without visible injury. This symptom has been reported from a variety of Jurassic and Cretaceous Belemnites and possibly results from a parasitic infestation This infestation caused the formation of a collar that is regarded as an effluence channel and potentially indicates an inflammation of the mantle sac epithelium. Hoffmann et al. report collar formation in one specimen of Belemnellocamax sp.
Most studies about modern Molluscan parasites deal with parasites of Bivalves. Bivalves host a diverse group of parasites and disease-causing agents such as, e.g., Viruses, Prokaryotes, Fungi, Protists, Parazoans, and Metazoans (Platyhelminths, Annelids, Molluscs, Bryozoans, and Arthropods). The most abundant parasites are endoparasitic Flukes (Trematoda, Digenea), which have also been observed in Gastropods. Larvae of Digenean Trematodes (Gymnophallidae) sometimes lead to the formation of blisters and have been found in fossil Bivalves dating back to the Triassic. Blister formation, however, can have a variety of causes, some of which are not due to parasites. Identification of a specific parasite taxon, however, is hampered by the fact that parasites are typically small-bodied (submillimetre) and usually lack biomineralised hardparts.
In modern Cephalopods, parasites are known from many species from all major oceans ranging from coastal, shelf, to oceanic and deep-sea environments. This is no surprise given that Cephalopods are a key trophic element in marine ecosystems. One study documented 200 species of endoparasites in Cephalopods with macroparasites such as Nematodes, Copepods, and Isopods. These predominantly affect non-mineralized tissues such as the digestive tract and the gills. The infestation of mainly non-mineralised tissues limits the fossil preservation potential of parasite activity and the resulting host reaction.
For Belemnites, most reported pathologies are due to survived predator attacks (e.g. bent rostra), a few malformations are assigned to the activity of parasites like, blister malformation, collar formation, and apex malformation. Although Hoffmann et al. identify a parasite infection as the most likely trigger of these pathologies, attributing the deformations to a specific parasite taxon is difficult to impossible. While some parasites leave characteristic patterns on their host, this is not necessarily definitive evidence of their presence. Different parasites can leave similar traces and taxonomically distant parasites can inflict similar symptoms on their hosts because of convergence in the evolution of host-exploitation strategies. Differential diagnosis for palaeopathological phenomena may lead to the conclusion that two or more alternative conditions remain as potential triggers.
Hoffmann et al.'s high-resolution computed-tomography-data represent an innovative and non-destructive way to analyse the internal structure of Belemnite rostra. computed-tomography-images revealed internal structures that allow the examination of a variety of pathological phenomena and to differentiate between different types of pathology. Most importantly, the computed-tomography-data allow to infer potential causes of these deformations (predator attack or parasite activity), therefore going beyond a classification of pathologies from the surface of a specimen alone. Hoffmann et al. thereby demonstrated the usefulness and feasibility of the application of non-invasive imaging methods to the field of palaeopathology. The data gained can be further used for the identification of various diagenetic alteration features in Belemnite rostra. computed-tomography-data turned out to perform better than magnetic resonance imaging-data in terms of spatial resolution and contrast.
See also...






Follow Sciency Thoughts on Facebook.