
The South American Paleogene vertebrate record is plethoric of well-known Mammalian groups, which enclose several peculiar forms. Of these, the order Notoungulata stands as one of the most prolific, diverse, and geographically widespread clades. During the Palaeogene, the Notoungulates reached a high degree of taxonomic diversity, although the basal radiations of the group show a relatively low morphological disparity, sometimes hindering a clear assessment of intra order affinities.
Two monophyletic major groups are usually recognized within Notoungulata: Toxodontia and Typotheria. Nevertheless, the basal relationships of the order are controversial, as some Palaeogene taxa fall in basal polytomies outside these groups in integrative phylogenetic hypotheses. The vast majority of these phylogenetic studies are based on cranial and dental characters. Conversely, despite postcranial remains also provide phylogenetic information, they were only included in a few analyses, which received relatively little attention. More recently, they started to be more frequently used for complementing cranial and dental character matrice.
Among postcranial elements, the calcaneum and astragalus (i.e., proximal tarsals) provide valuable and highly useful information at systematic and morphofunctional levels. They are compact bones of the hind limb that form the joint between the zeugopodium and autopodium, and thus their importance as a functional unit is related to the locomotor adaptations of the studied Animals. Also, since each element functions as an integrated part of the hind limb, it is generally stable, as any modification of one element changes its role in relation to the remaining pieces of the functional complex. Consequently, their morphology can bear valuable information about the group they belong to, spurring an increasing interest in studies of these elements.
More interestingly, the calcaneum and astragalus are frequently found in the fossil record, only outcompeted by the teeth, which are usually the most common remains recovered. The Palaeogene levels from northwestern Argentina are not an exception to that pattern. In late Eocene outcrops of the Geste Formation of Catamarca Province, a 2016 study reported several proximal tarsals assigned to early-diverging Notoungulate taxa and archaic members of several lineages of South American native Ungulates. Moreover, middle–late middle Eocene sequences, recognised as the Lower and Upper Lumbrera formations of Salta Province, respectively, yielded several Vertebrate faunas. Within these faunas, Notoungulates are very frequent, represented by well-preserved, mostly cranial remains, often associated with postcranial elements. Particularly, Typotheria taxa have been described in detail, but all of the studies focused on cranial and dental elements.
In a paper published in the journal Acta Palaeontologica Polonica on 23 March 2020, Matías Armella of the Cátedra de Paleontología and Instituto de Estratigrafía y Geología Sedimentaria Global at the Consejo Nacional de Investigaciones Científicas y Técnicas, the Facultad de Ciencias Naturales e Instituto Miguel Lillo at the Universidad Nacional de Tucumán, the Facultad de Ciencias Exactas y Naturales at the Universidad Nacional de Catamarca, and the Instituto Superior de Correlación Geológica at the Consejo Nacional de Investigaciones Científicas y Técnicas, Daniel García-López also of the Cátedra de Paleontología at the Universidad Nacional de Tucumán, the Instituto Superior de Correlación Geológica at the Consejo Nacional de Investigaciones Científicas y Técnicas, Judith Babot also of the Fundación Miguel Lillo at the Consejo Nacional de Investigaciones Científicas y Técnicas, Virginia Deraco, also of the the Cátedra de Paleontología at the Universidad Nacional de Tucumán, the Instituto Superior de Correlación Geológica at the Consejo Nacional de Investigaciones Científicas y Técnicas, Claudia Herrera and Luis Saade also of the Cátedra de Paleontología at the Universidad Nacional de Tucumán, and Sara Bertelli of the Unidad Ejecutora Lillo at the Consejo Nacional de Investigaciones Científicas y Técnicas, and the Fundación Miguel Lillo, analyse anatomical and systematic aspects for tarsals of notoungulates recovered from the Lower and Upper Lumbrera formations. Most of these elements are associated with cranial remains and are described here for the first time; the fact that they are well preserved and articulated allows us to undertake a detailed analysis. Finally, they consider several morphological features of the tarsals to infer Mammalian foot stances and range of movements. Given that the Palaeogene is considered a period of major importance for the evolution and diversification of Notoungulata, Armella et al.'s contribution enhances the tarsal morphology as a trait to assess phylogenetic affinities and morphofunctional patterns in early forms within Typotheria.

The first specimen examined is PVL 6227, the right calcaneum and left and right astragal assigned to the Typotherian Notoungulate Colbertia lumbrerense, part of a skeleton of an adult individual also preserving the skull and mandibles, from the middle Eocene (Vacan subage) Lower Lumbrera Formation of Pampa Grande in Guachipas Department, Salta Province, Argentina.
The calcaneum is poorly preserved. This element has suffered a dorsoplantar compression that altered its main features and it will not be described here. The astragali are complete, but the right one is also compressed (dorsoplantarly); the following description is based on the better-preserved left element. In dorsal view, the tibial trochlea is shallow, asymmetric, and trapezoidal in shape, the lateral crest being more developed and conspicuous than the medial crest, which is more rounded. The trochlear fossa is shallow and almost smooth. The astragalar neck is short compared to other Paleogene taxa (e.g., Thomashuxleya externa, Notopithecus adapinus) and other specimens studied by Armella et al., representing almost 28% of the total length of the astragalar body. The oblique dorsal crest (nuchal crest or tibial stop) is robust, oriented posterolateral-anteromedially and it occupies the entire surface of the neck (it forms an angle of approximately 62° with the anteroposterior axis of the astragalus). The head is slightly eroded on its medial side; however, it can be observed that it is spherical, being roughly as wide as the neck but not expanded (a shallow groove surrounding the head separates both structures). The navicular facet occupies the entire anterior surface of the astragalar head and it is slightly extended on the medial surface.

In plantar view, the astragalus shows some eroded areas; nevertheless, the articular facets are well-preserved. The ectal facet is subtrapezoidal, with the major axis anteroposteriorly oriented. It is represented by a deep concavity that faces lateroplantarly. In contrast, the sustentacular facet is convex, oval, and it occupies the plantar surface of the astragalar neck. This facet exhibits well-defined edges. Although a small, smooth surface connects the sustentacular and navicular facets, the strong concavity of this surface indicates that both facets are functionally separated.
The interarticular sulcus is deep and anterolateral-posteromedially oriented. Moreover, in its posterior area, the inferior astragalar foramen is small and faces anteroplantarly. The groove for the tendon of the flexor hallucis longus muscle is poorly developed compared to other specimens such as PVL 4292 and it is roughly continuous with the trochlea. The lateral crest that limits the groove is broken; nevertheless, the rest of the structure is clearly visible. The dorsal astragalar foramen is present but very small and located near the lateral crest of the trochlea. On the medial area, the astragalar medial plantar tuberosity is small but well pronounced, defined by the anteromedial portion of the medial crest. The orientation of the facet for the medial malleolus of the tibia is similar to the astragalar medial plantar tuberosity, but the former is smooth and less projected. The attachment for the tibioastragalar ligament is located between these two structures, forming a shallow groove.
In lateral view, the fibular facet is flat, crescent-like (with a blunter, although somewhat eroded, posterior end), and lacking projections. The attachment for the fibuloastragalar ligament is represented by a deep and conspicuous kidney-shaped depression, with the major axis being anteroposteriorly oriented. In medial view, the medial aspect of the astragalar medial plantar tuberosity is elliptical, with the major axis being anteroposteriorly oriented. Although there is an eroded area on the medial surface, this structure is not connected with the navicular facet (this is more evident on the right astragalus).
The genus Colbertia includes two species of small-sized Notoungulates: Colbertia magellanica, from the early Eocene of Brazil (Itaboraí) and Colbertia lumbrerense, recorded from middle Eocene rocks of the Lower Lumbrera Formation in Salta Province, Argentina. Although previous works have studied postcranial remains of both species, these authors did not provide detailed descriptions of the tarsal bone morphology, since these elements were not fully prepared and poorly preserved in the specimens available to them. In the particular case of Colbertia lumbrerense, one specimen analysed in a 2007 was the postcranium PVL 6218, currently not available to us, and proximal tarsals were not figured in detail.
The recently prepared PVL 6227 allows Armella et al. to compare in detail the two species of Colbertia. Concerning the astragalus, the shared features between Colbertia magellanica and Colbertia lumbrerense are the spherical astragalar head, where the sustentacular and navicular facets are separated by a deep sulcus, and the navicular facet slightly extended on the medial surface, which is not connected with the astragalar medial plantar tuberosity.
It was considered that, at least regarding the features exposed in the specimen PVL 6218, the trochlea and ectal facet morphology are traits shared by Colbertia magellanica and Colbertia lumbrerense. Nevertheless, PVL 6227 shows an asymmetric and trapezoidal tibial trochlea, in contrast to Colbertia magellanica (MCT 2456M), where the trochlear crests are parallel and inclined to the lateral side are the ectal and fibular facets not projected laterally, smaller astragalar medial plantar tuberosity on the medial face, longer neck (made of the oblong sustentacular facet), and very small dorsal astragalar foramen (in Colbertia magellanica this foramen is well developed and located in a depression on the trochlea). In summary, the mentioned differences are opposed to a previous point of view that considers the morphologies of Colbertia magellanica and Colbertia lumbrerense being similar. Hence, these comparisons indicate a source of diagnostic traits potentially useful to the further characterization of both species.
The second specimen examined is PVL 4292; left calcaneum, astragalus, and associated distal fragment of the fibula form an indeterminate Typotherian Nonounguate; from Pampa Grande, Guachipas Department, Salta Province, Argentina Lower Lumbrera Formation, middle Eocene.
The elements are well preserved and the articular facets can be clearly observed. Nevertheless, there are broken areas and surfaces that have been eroded, as evidenced by angular edges (e.g., the middle region of the calcaneum and the astragalus neck).
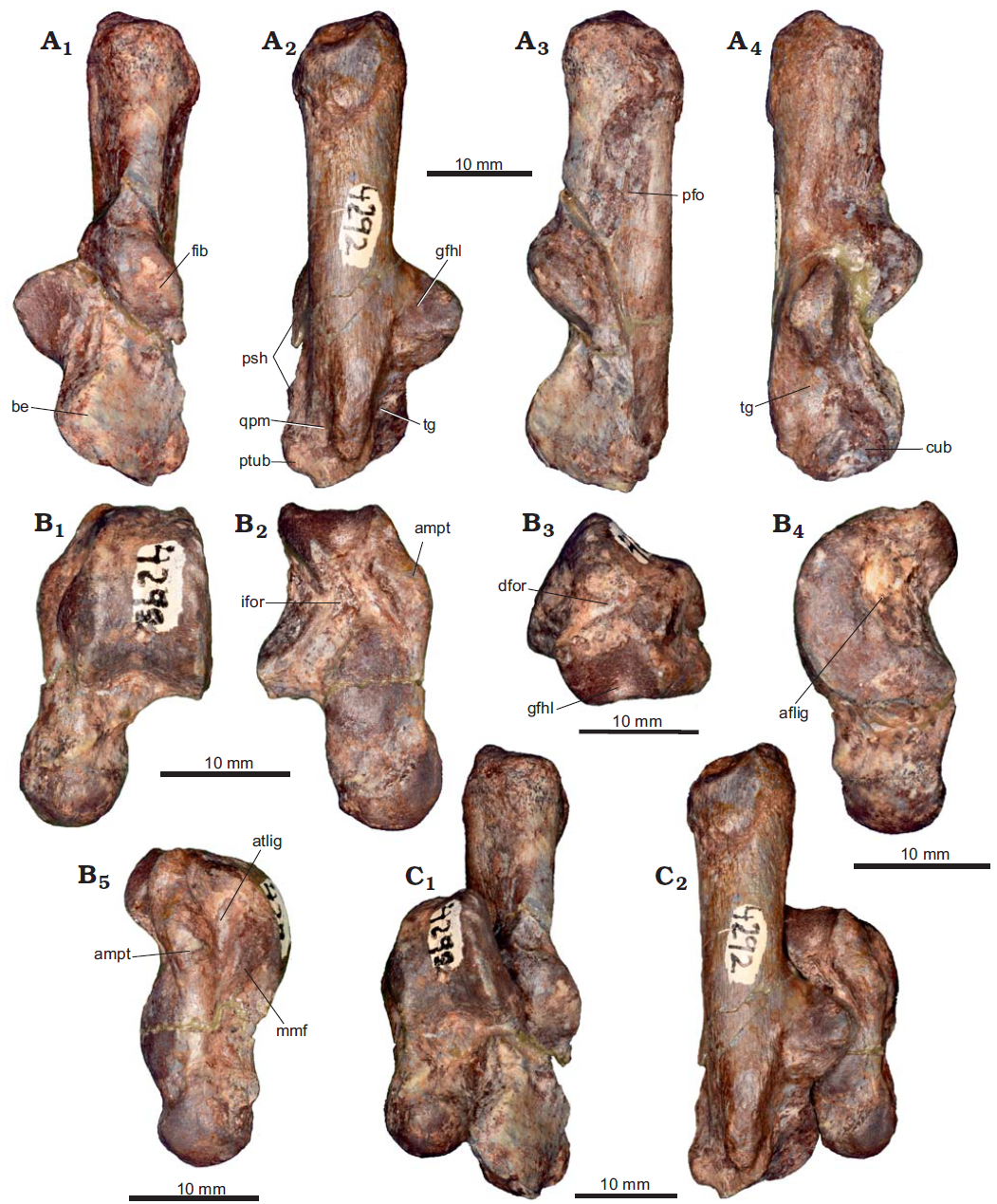
The Calcaneum is an elongated and relatively robust bone, compared with the other specimens here studied. In dorsal view, the tuber calcis represents almost 45% of the calcaneum total length. The dorsal surface is narrower than the plantar face; hence, the cross section of the tuber is subtriangular. The apical region is also wider than the dorsal aspect of the tuber and its posterior surface is rugose and dorsoplantarly sloping. On the tuber neck, a small concavity can be observed posteriorly and laterally to the fibular and ectal facets, respectively. The ectal facet is convex and medially oriented; it is crescent-shaped with a wider posterior area. Adjacent to this, the fibular facet is also markedly convex but it is larger (in comparison to other Eocene ungulates; see below) and anterodorsally oriented. It is subrectangular with markedly rounded edges, with the major axis obliquely arranged compared to the major axis of the calcaneum. Both ectal and fibular facets form a conspicuous ectal protuberance. The sustentaculum is larger than the ectal protuberance and anteromedially located. The sustentacular facet is smaller than the ectal and fibular facets. It is dorsally oriented, slightly concave, and subcircular; however, its anterior edge is slightly projected. Between the sustentacular and ectal facets, there is a broken area which prevents the assessment of the dimensions and morphology of the interarticular sulcus. In the anteromedial end of the calcaneum and separated from the sustentacular facet, the anterior astragalocalcaneal facet is slightly elongated on a large dorsal projection.
In plantar view, the surface for the attachment of the muscle flexor hallucis brevis has rounded edges and occupies the entire width of the tuber apex. The groove for the tendon of the flexor hallucis longus muscle is broad and limited by a shallow longitudinal crest on the plantar aspect (tendinous groove). Toward the anterior region, the most prominent feature is the plantar tubercle. It is a robust structure that is developed parallel to the major axis of the calcaneum for the plantar ligaments (to the cuboid and cuneiforms) and tarsal fibrocartilage.
On the lateral side, the peroneal fossa is bounded by the ectal protuberance and two longitudinal crests. This fossa occupies almost 42% of the calcaneum total length and it is much longer than the anteroposterior extension of the fibular facet. The peroneal tubercle is a robust process located on the anterior end of the lateral face. Despite being posteriorly broken, it seems to be extended from the anterior edge of the ectal protuberance to the anterolateral end of the calcaneum, forming a peroneal shelf. Anterolaterally, on the peroneal shelf, a well-marked groove for the attachment of the quadratus plantae muscle is more exposed in plantar view.
In medial view a tendinous groove is located medial to the plantar tubercle and separated from the anterior edge of the sustentaculum. It is slightly concave, oblique, and does not extend to the anterior face. According to one 1998 study, this is a site for attachment of calcaneocuboid ligaments. The calcaneum cuboidal facet is clearly observed in this view; it is strongly concave and has an elliptical shape with an anteroplantar to posterodorsally oriented major axis, reaching almost a complete medial exposure.
The astragalus is more robust than the calcaneum and represents 53% of the calcaneum total length. In dorsal view, the tibial trochlea is asymmetric, trapezoidal, and has a slightly excavated trochlear concavity. The lateral trochlear crest is wider than the medial crest, which shows more rounded edges. The trochlear concavity leads anteriorly to a moderately deep trochlear fossa, which is in turn bounded by the anterior end of the lateral trochlear crest. The oblique dorsal crest is well defined and developed in a posterolateral-anteromedial direction; it forms an angle of approximately 58° with the anteroposterior axis of the astragalus. The astragalar neck is longer and narrower than the astragalar body; it is slightly oblique, causing a minimum medial displacement of the astragalar head. The head is spherical and almost as wide as the neck; both areas are separated by a shallow groove. The navicular facet extends on the medial surface of the neck.
In plantar view, the ectal facet is well defined; it is sub-triangular with a vertex pointing anterolaterally. This facet is strongly concave and faces laterally. In contrast, the sustentacular facet is slightly convex, teardrop-shaped, and occupies a large portion of the astragalar neck. This facet exhibits well-defined edges, and it is separated from the navicular facet by a deep sulcus. The articular facet arrangement of the proximal tarsals PVL 4292, results in a transverse articulation plane (side by side), rather than an overlapping disposition of both elements. Between the ectal and sustentacular facets, the interarticular sulcus is deep, anterolateral-posteromedially oriented, and it leads to a small inferior astragalar foramen. In the posterior area, the groove for the tendon of the flexor hallucis longus muscle is wide and defined by two sharp crests. This groove is not continuous with the tibial trochlea. With the astragalus and calcaneum in articulation, the groove is continuous with a similar structure on the calcaneum, completing the passage for the flexor hallucis longus muscle. There is a small notch or fossa developed on the surface separating the trochlea and flexor groove, clearly visible in posterior view. This structure probably houses the dorsal astragalar foramen; however, the bottom is filled with sediment matrix and the presence of an aperture is not certain. If present, the foramen would be very small.
In lateral view, the fibular facet and the attachment for the fibuloastragalar ligament are similar to those observed in Colbertia lumbrerense (PVL 6227). On the medial side, the astragalar medial plantar tuberosity is larger and wider than the facet for the medial malleolus of the tibia. It is fusiform in medial view, with the major axis anteroposteriorly oriented. The facet for the medial malleolus of the tibia is adjacent to the aforementioned tuberosity and presents almost the same development and orientation. Both structures are separated by the attachment for the tibioastragalar ligament. It is a deep groove, wider on the posterior region and narrower toward the anterior area.
Previous studies recognised several synapomorphies in the astragalus of Notoungulata: astragalar neck constricted and well-differentiated from the head and the body, presence of the oblique dorsal crest, marked astragalar medial plantar tuberosity, and sulcus extended laterally from the dorsal astragalar foramen (generally absent in derived Notoungulates). Moreover, on 1998 study added the medially extended navicular facet as a diagnostic feature for the clade. The presence of this set of distinctive traits results in the identification of PVL 4292 as a Notoungulate.
Several notoungulate tarsals are known for the Palaeogene; however, only a few records came from Eocene levels. In this context, PVL 4292 was initially compared with specimens sharing temporal affinities.
The basal notoungulate Notostylops murinus (Notostylopidae) was reported for the middle Eocene of Chubut, Argentina. Its astragali (MACN-A 10940) differ from PVL 4292 in parallel trochlear crests, laterally inclined; short astragalar neck, narrower than the head; small astragalar medial plantar tuberosity; lateral process well developed; ectal facet more plantarly exposed; and a large dorsal astragalar foramen. Regarding Allalmeia atalaensis, another basal form found in middle Eocene levels of Mendoza, Argentina, several features distinguish it from PVL 4292. Although the calcaneum of Allalmeia atalaensis (MCNAM-PV 507) is poorly preserved, it shows a more circular sustentaculum, well-developed peroneal tubercle, and a more horizontal orientation of the cuboidal facet. Besides this, the astragalus MCNAM-PV 507 differs from PVL 4292 in the presence of well-developed lateral process and astragalar medial plantar tuberosity, and small facet for the medial malleolus of the tibia.
The Eocene Toxodontians that preserve the proximal tarsals are the genera Thomashuxleya and Periphragnis ('Isotemnidae'). Regarding the calcaneum, only the PVL 316 (Thomashuxleya sp.) was available for comparison; it is almost two times larger than PVL 4292 and shows a robust tuber calcis with a globose posterior region. Also, the fibular facet is very small, the ectal facet is large (pointing to the lateral and anterior faces), and the cuboidal facet is more transversely oriented than in PVL 4292. As for the astragalus, both Thomashuxleya (PVL 332) and Periphragnis differ from PVL 4292 in the presence of a wide trochlea, large dorsal astragalar foramen, well-projected astragalar medial plantar tuberosity, short neck, and a shallow oblique dorsal crest. Additionally, Bruce Shockey and John Flynn described another middle Eocene indeterminate Isotemnid from Patagonia, Argentina, which preserves an almost complete foot. In overall terms, this specimen (AMNH 28690) also displays the differences mentioned above, except for the ectal facet, which is more plantarly oriented in the astragalus, and the tuber calcis, which is dorsoplantarly wider than in PVL 4292.
Within Typotheria, the astragalus of PVL 4292 differs from the Oldfieldthomasiid Colbertia lumbrerense (PVL 6227) in several traits: larger size, which is 46% longer than PVL 6227; presence of a longer neck; narrow trochlea compared to the ratio between total length of the astragalus and width of the trochlea; ectal facet exposed mostly on the lateral side; and presence of a notch or fossa housing the dorsal astragalar foramen (although this last character is not fully visible; see above). Regarding Colbertia magellanica, the same differences are observed, with the remarkable exception of the notch for the dorsal astragalar foramen, present also in this species. As for the calcaneum (which cannot be compared with Colbertia lumbrerense) PVL 4292 differs from Colbertia magellanica (MCT 1263M) in a more vertically oriented ectal facet; well- developed fibular facet; and a subcircular sustentacular facet.
The tarsal morphology is also known for Eocene interatheriids. Bárbara Vera described the postcranial morphology of Notopithecus adapinus, from the middle Eocene of the Argentinean Patagonia. Although the tarsals show a few similarities (e.g., well-developed peroneal fossa, one concavity near the ectal prominence in the calcaneum, asymmetric, shallow, and narrow astragalar trochlea, and a long neck with spherical head in the astragalus), PVL 4292 differs from this specimen (MPEF-PV 1113) in a longer tuber calcis compared to the calcaneal body; low development of the peroneal shelf in the calcaneum; and a larger astragalar medial plantar tuberosity and greater development of the medial trochlear crest in the astragalus. In addition, the PVL 4292 remains are almost four times larger than the tarsals of Notopithecus.
In northwest Argentina, the Eocene Geste Formation yielded several proximal tarsals that were assigned to Interatheriidae (two calcanei and one astragalus). Morphologically, the calcanei MHAS 046 and MHAS 047 share with PVL 4292 a long tuber calcis; wide peroneal fossa; fibular facet markedly convex and obliquely arranged regarding the major axis of the calcaneum; projected ectal protuberance; small and slightly concave sustentacular facet with its anterior edge slightly projected; presence of a small concavity posteriorly and next to the fibular and ectal facets, respectively; and robust plantar tubercle. Furthermore, although the astragalus MHAS 048 lacks the lateral crest of the trochlea, it shows a long neck and spherical astragalar head similar to the astragalus PVL 4292. However, the discrepancy in size between these tarsals and the specimen PVL 4292 is remarkable and reaches the same values as that mentioned for Notopithecus adapinus.
At this point, a recent taxonomic proposal made by Bárbara Vera should be considered. This author performed a cladistic analysis including both cranial and postcranial characters and proposed a drastic change in the systematic of the forms usually considered as basal interatheriids. In this contribution, Notopithecus, Transpithecus, Antepithecus, and Guilielmoscotia are recovered as a monophyletic clade named Notopithecidae and defined by two unequivocal synapomorphies: small mesial extension of the entolophid and asymmetrical astragalar trochlea with higher lateral side more oblique than the medial side. Considering these traits, the specimen PVL 4292 should be included as a member of Notopithecidae. Moreover, other general characters are mentioned for the group and are present in the material studied here: inclined fibular facet, oblique astragalar medial plantar tuberosity, and absence of astragalar-cuboid contact. Nevertheless, in addition to the already mentioned differences with Notopithecus (which include a much larger size), it should be noted that postcranial traits are only known for this latter genus within the focus group studied by Bárbara Vera, and that this particular morphology for the astragalar trochlea is widely distributed among other Palaeogene Notoungulates (e.g. Allalmeia, Colbertia, 'Campanorco', Trachytherus). It is clear that this taxonomic proposal should be evaluated in light of new materials and may be challenged in future analyses, and thus Armella et al. follow here a more traditional taxonomic approach.
It is also important to mention that several of the aforementioned features are shared with interatheriids recorded from the late Oligocene (e.g. Federicoanaya) and the middle Miocene (e.g. Protypotherium). Additionally, these post-Eocene taxa show a well-defined facet in the calcaneum for the articulation with the navicular. The last trait is also observed in some Toxodontians (e.g. Leontiniids, Notohippids, and early Toxodontids), but it is absent in PVL 4292.
In summary, there is a set of similarities that would indicate that the specimen PVL 4292 is close to basal Interatheriids (in a traditional taxonomic scheme). Nevertheless, it is clear that the remarkable larger size of the former represents an important difference compared to any basal Interatheriid taxon, which show mostly small body sizes. In addition, although the Lower and Upper Lumbrera formations levels are highly fossiliferous, there is so far no record of that family therein. A direct assignment is thus difficult, as it would imply the presence of a very large, and thus rather unusual, basal interatheriid morphotype in these levels. Armella et al. therefore refrain from referring PVL 4292 to this clade, provisionally favoring its identification as a basal Typotheria.
The name 'Campanorco inauguralis' was introduced in 1984 in one of the abstracts of a scientific meeting. Although in this abstract even a family name was also included (Campanorcidae) no formal definition or illustration was provided; hence, the erection of this taxonomical entity does not fit in the standards of the International Commission of Zoological Nomenclature. Nevertheless, this term has been widely used in literature, sometimes regarded as an 'informal taxon'.
Additionally, a formal description of the first material mentioned as 'Campanorco' and other new specimens is currently under way, dealing also with the proper use of this term. Thus, here we maintain the use of both the generic and specific epithets between quotation marks in order to link our observations with previous contributions and to avoid further confusion until this nomenclatural issue is resolved.
IBIGEO-P 105 is assigned to 'Campanorco inauguralis'. This is articular portion of left calcaneum, almost complete right calcaneum, complete left astragalus, trochlear portion of right astragalus, and left cuboid and navicular. These elements are part of an incomplete skeleton including limbs, pelvis, vertebral column, and cranial remains; from El Simbolar, Guachipas Department, Salta Province, Argentina; Upper Lumbrera Formation, late middle Eocene (Barrancan subage of the Casamayoran).
The bones belong to a juvenile individual (this is inferred by the presence of deciduous dentition).
Both tuber calcanei are incomplete, preventing from estimating the total length of this element. In dorsal view, it seems to be more robust than the calcaneum PVL 4292; however, both present a plantar surface that is wider than the dorsal surface. The tuber neck has a small concavity on the posterior region of the ectal protuberance. The ectal facet is convex (although less than in the calcaneum PVL 4292), elliptical in shape, and medially oriented. The fibular facet is also convex; it is more slender than in PVL 4292 but shows the same arrangement. Both facets are better observed in the right calcaneum, where they are differentiated by a change of orientation of the surface. The sustentaculum is well developed medially and the sustentacular facet is larger than the fibular facet but subequal in size to the ectal facet. The sustentacular facet is also dorsally oriented, slightly concave, and circular with crested and well-defined edges. In the anteromedial end of the calcaneum, the astragalocalcaneal facet is small and well separated from the sustentacular facet.

In plantar view, the groove for the tendon of the flexor hallucis longus muscle is narrower and shallower than in the calcaneum PVL 4292. The plantar tubercle is slightly projected although developed on the anterior surface of the element; it is smaller than in the PVL 4292; however, this could have been affected by erosion.
On the lateral aspect, the peroneal fossa is poorly developed and located above the level of the ectal protuberance. This condition is opposite to that seen in the calcaneum PVL 4292. The peroneal tubercle is weak and rugose; it does not form a peroneal shelf. In medial view, the tendinous groove is slightly concave and restricted to a small area, medial to the plantar tubercle and anteroplantar to the navicular and sustentacular facets. The cuboidal facet is strongly concave and elliptical with the major axis anteroplantar to posterodorsally oriented. It is smaller and less inclined than in the calcaneum PVL 4292.
A remarkable feature of these calcanei, clearly visible on the right one, is the presence of a facet for the navicular bone. It is evident on medial view, between the anterior region of the sustentaculum and the cuboidal facet, and it is much smaller than the sustentacular facet, slightly concave, and elliptical.
This bone is somewhat flattened dorsoplantarly; despite this, it can be observed that this is mid-sized regarding the other astragali reported by Armella et al. It is 23% smaller than the astragalus PVL 4292 and 28% larger than the astragalus of Colbertia lumbrerense (PVL 6227). In dorsal view, the tibial trochlea is slightly asymmetric, trapezoidal, and more excavated than in the astragali described. The medial crest is well developed, reaching the medial side of the astragalar neck (a somewhat similar condition is present in Colbertia lumbrerense, although in this case the anterior end of the crest is weaker). The trochlear fossa is deeper than in the astragalus of Colbertia lumbrerense, but shallower than in PVL 4292. Anterior to this fossa, the oblique dorsal crest is conspicuous but short; it forms an angle of approximately 45° with the anteroposterior axis of the astragalus. Unlike other notoungulate astragali this crest is confined to the lateral area of the astragalar neck. This neck is slightly narrower than the head, in which the navicular facet occupies the entire anterior extension. The main exposure of this facet is dorsal; in plantar view it is almost completely hidden. Moreover, it extends on the medial surface of the astragalar neck. Such a condition was observed in the other astragali described by Armella et al. The astragalar head seems to be more developed transversely. However, this condition is related to dorsoplantar deformation, since the astragalar facet of the navicular bone (in which the head articulates) is markedly concave and hemispherical.
In plantar view, the arrangement of the articular facets of IBIGEO-P 105 is similar to that in Colbertia lumbrerense. Nevertheless, the ectal facet is proportionally larger and more laterally extended than in the latter. Furthermore, the sustentacular facet is subcircular, with well-defined medial and lateral crested edges, and it is well separated from the navicular facet by a deep sulcus. The sustentacular facet is larger than in Colbertia lumbrerense. Between the ectal and sustentacular facets, the interarticular sulcus houses the inferior astragalar foramen, which is posteriorly located. As in Colbertia lumbrerense (PVL 6227), the groove for the tendon of the flexor hallucis longus muscle is not well developed. The dorsal astragalus foramen is large and located on the surface between the astragalar trochlea and flexor groove (interrupting the contact between both structures). Although the edges of this foramen are broken, it seems to have been rounded. This condition is different from the astragalus PVL 4292, in which it is located into a notch, and from Colbertia lumbrerense, in which this foramen is very small and more dorsolaterally placed. On the medial area, the astragalar medial plantar tuberosity is projected medially.
In lateral view, the fibular facet is concave and crescent- like, with the major axis anteroposteriorly and obliquely oriented. Moreover, the lateral process is prominent, a feature not observed in the astragali described above. The attachment for the fibuloastragalar ligament is shallower and proportionally smaller than in Colbertia lumbrerense and the astragalus PVL 4292. On the medial aspect, the astragalar medial plantar tuberosity is elliptical, with the major axis anteroposteriorly oriented, and it does not reach the astragalar neck. The facet for the medial malleolus of the tibia is smaller than the astragalar medial plantar tuberosity, and both structures are separated by a slight depression corresponding to the attachment for the tibioastragalar ligament.
The navicular bone is not affected by deformation. It only preserves the corpus, which is robust and anteroposteriorly short. In posterior view, there is a facet for the astragalar head occupying the entire surface. It is very concave and circular. On the anterior surface, there are three main facets and a prominent tuberosity. The facet located on the lateral end is for the cuboidal bone; it forms a right angle with a small lateral facet for the calcaneum. The cuboidal facet is elliptical, slightly convex, and faces medially. Between this and the other facets, there is a deep sulcus dorsolaterally to medioplantarly oriented. The remaining facets are separated from each other by a shallow crest. The medial facet (which is larger) articulates with the mesocuneiform, while that for the ectocuneiform is lateral. The mesocuneiform facet is subtriangular and concave-convex, whereas the ectocuneiform facet is teardrop-shaped and slightly convex. There is no articulation for the entocuneiform. On the plantar region, there is a robust tuberosity that would be connected with the plantar apophysis. Finally, there is a triangular broken surface on the medial aspect of the navicular. This area may have corresponded to the navicular medial tuberosity present in basal Notoungulates. Nevertheless, this cannot be ascertained.
The cuboid is well preserved. The corpus is robust and trapezoidal. The posterior surface is occupied by the facet for the calcaneum, better observed in dorsal view; it is strongly convex and mediolaterally inclined. Medially, there are two facets separated by a deep sulcus. The posterior facet is that for the navicular and the anterior facet represents the articulation for the ectocuneiform. The navicular facet is almost elliptical and larger than the ectocuneiform facet, which is dorsoplantarly elongated and seems to be formed by two lobes with a smooth concavity between them. In anterior view, the metatarsal facet is concave and teardrop-shaped. On the plantar side there is a deep sulcus running mediolaterally, which houses the tendon of the peroneous longus muscle. The posterior edge of this sulcus is formed by a robust tuberosity that occupies almost the entire plantar surface of the cuboid.
'Campanorco inauguralis' is one of the most frequent Notoungulates from the Upper Lumbrera Formation. Even though it was only informally reported in nomenclatural terms, it was included in several phylogenetic analyses within Notoungulata. Initially 'Campanorco inauguralis' was considered to be close to Mesotheriidae, based on several dental and cranial features. Later, it was grouped into Typotherioidea, along with Mesotheriidae, Hegethotheridae, and polyphyletic Archaeohyracidae. In turn, postcranial remains of 'Campanorco inauguralis' were unknown so far, and therefore their description could contribute to assessing these proposed phylogenetic affinities.
Among basal notoungulates, the astragali of Notostylops murinus (MACN-A 10940) show a similar size regarding 'Campanorco inauguralis'; nevertheless, these bones differ in the marked asymmetrical trochlea with parallel and laterally inclined crests, and a lateral process strongly projected. The basal typotherian Colbertia lumbrerense (PVL 6227) differs from IBIGEO-P 105 in a small-sized astragalus, an astragalar medial plantar tuberosity longitudinally extended, and a small lateral process. Additionally, Colbertia magellanica (MCT 2456M) shows a short and robust astragalar neck and more developed astragalar medial plantar tuberosity and lateral process than in 'Campanorco inauguralis'. The calcaneum of Colbertia magellanica (MCT 1263M) is also markedly different in shape and the arrangement of the articular facets (e.g., ovoid sustentacular facet, ectal facet more obliquely oriented) compared to 'Campanorco inauguralis'.
Concerning Interatheriidae, recovered as the sister clade of Typotherioidea, Notopithecus adapinus (MPEF-PV 1113) displays a slender and small astragalus, with a markedly asymmetrical trochlea. The calcaneum of Notopithecus adapinus differs from 'Campanorco inauguralis' in the presence of a robust tuber calcis, a low inclination and development of the cuboid facet, and a small and oblique fibular facet. In addition, interatheriid calcanei from the Geste Formation present two concavities adjacent to the ectal protuberance, while there is one concavity in 'Campanorco inauguralis', and they have an ectal facet that is less vertical.
Finally, comparisons among the known postcranial remains for basal Mesotheriids are crucial due to the aforementioned hypothesis relating this family with 'Campanorco inauguralis'. In this sense, Paleogene tarsals are known for Trachytherus alloxus and Trachytherus ramirezi, from the late Oligocene of Salla, Bolivia. The astragali of these taxa (UF 172437 and MUSM 961) differ from 'Campanorco inauguralis' in the presence of a markedly asymmetrical trochlea, short neck, and small head in relation to the astragalar body. In addition, the calcaneum UF 172514 (Trachytherus alloxus) shows a small sustentaculum, an oblique orientation of the ectal facet, lesser development of the fibular facet, and an almost transverse cuboid facet. Furthermore, in derived Mesotheriids such as Mesotherium the mentioned features do not show any significant modification in the observed tarsals (i.e., MACN-PV 1950, 1975, 7760) compared basal representatives of the family and, therefore, the morphology is also different from 'Campanorco inauguralis'.
One of the most remarkable features in the calcaneum of 'Campanorco inauguralis' is the presence of a facet for the navicular bone. This was considered by Richard Cifelli as a 'reversed alternating tarsus' condition, where the astragalus lacks its contact with the cuboid while the calcaneum articulates with the navicular bone. The calcaneal-navicular articulation was observed also in Leontiniidae, 'Notohippidae', Toxodontidae, and some Typotherians (e.g. Protypotherium and Federicoanaya), as well as in Lagomorphs and Arctostylopids. This fact, added to the differences observed with the compared mesotheriids (i.e. Trachytherus and Mesotherium), such as a reduced fibular-calcaneal contact, oblique ectal facet of the calcaneum, and the astragalus overlying the calcaneum, among others, highlight the singular condition of the proximal tarsals of 'Campanorco inauguralis'.
Based on the comparative analysis and the combination of features exhibited by 'Campanorco inauguralis', the proximal tarsals contain useful information for diagnostic purposes. In turn, there are no characters to support the hypothesis of phylogenetic affinities between this form and the Mesotheriidae clade, and the specimen analysed by Armella et al. shows a generalized morphology within Notoungulata, considering both basal and derived taxa. This is in agreement with previous hypotheses that locate Campanorcids in an unclear and basal position within Typotheria.
The significance of tarsals for palaeobiology stems from the fact that they are usually in direct contact with the substrate; thereby, these elements are immediately related to locomotor habits and foot postures. Even though the articulation between calcaneum and astragalus is usually a relatively static joint, this assemblage works as a hinge between the tibia-fibula and the foot bones. In most Mammals, the proximal tarsals articulate through a complex arrangement of contact facets. Given that, the morphology of the calcaneum and astragalus makes possible the inference of Mammalian foot posture and range of posterior limb movements.
The Mammalian foot can assume a range of postures: plantigrady, digitigrady, and unguligrady Both plantigrady and digitigrady are widespread conditions among basal terrestrial mammals, including several taxa among the South American native Ungulates, whereas unguligrady is a highly specialized condition associated with several morphological changes and restricted to more derived taxa.
Biomechanically, the calcaneum is the bone where the astragalus rests, functioning together, along with other elements, as a crurotarsal lever system with reference to the foot. In this model the calcaneum acts as a moment arm or in-lever, whereas the metatarsals represent the out-lever. The functional principle of these components is that a relatively longer in-lever increases the amount of out-force produced by a given in-force. In this context, the calcaneum in Typotheria indet. and 'Campanorco inauguralis' presents a long tuber calcis compared to total length, which would be effective in an animal that requires greater power in the foot stroke, but not velocity. This arrangement is common in plantigrade Mammals.
Concerning the astragalus, the trochlea articulates with the tibia and moves anteroposteriorly during locomotion. Likewise, the position of the astragalus on the calcaneum sets the plane of the major axis of the tibia. The three specimens analysed here (i.e. Colbertia lumbrerense, Typotheria indet., and 'Campanorco inauguralis') show an asymmetric trochlea with crests of different sizes (the lateral one is the largest). In this configuration, Matthew Carrano recognised that in plantigrade Mammals the tibia goes through trajectories of different radii over the two crests, and that it does not move in a strictly parasagittal plane. It tends to move along the smaller radius (i.e. the narrower crest), since if it moves along the larger radius (i.e. the trochlear concavity and greater crest) it would produce dislocation on the other radius. Added to this, a shallow trochlear concavity allows mediolateral movements of the foot, and thus, the resulting instability is offset by the open position of the phalanges, observed in plantigrade forms. In the case of a symmetric astragalar trochlea, the tibia describes an arc along the midpoint and rotates without significant mediolateral movement. This latter arrangement is important given the metatarsals-phalanges setting in digitigrade Mammals.

In the anterior region, the astragalar head is involved in foot rotation, which is associated with the relation among the mediolateral axis of the astragalar head, trochlear crests and metatarsal planes, and the major axis of the tibia. In Colbertia lumbrerense, Typotheria indet., and 'Campanorco inauguralis' the mediolateral axis of the astragalar head is parallel to the plane of the trochlear crests, so that the trochlear plane draws an angle with the metatarsal plane. As a result, when the metatarsals flex and extend, they do so at an oblique (rather than perpendicular) angle with respect to the major axis of the tibia. This arrangement is common in plantigrade forms, whereas in digitigrade Mammals the mediolateral axis of the astragalar head is oblique to the trochlear crests; thereby, the trochlear and metatarsal planes are parallel. Thus, the metatarsals rotate in a plane perpendicular to that of the long axis of the tibia in digitigrades.
Lastly, the presence of a dorsal astragalar foramen (frequent in plantigrade Mammals) is considered as a feature that limits dorsoplantar movements. The fact that this foramen is usually associated with a structure that interrupts the connection between the astragalar trochlea and the groove for the deep digital tendons supports this inference. Additionally, the development of a robust oblique dorsal crest on the astragalar neck also restricts the foot flexion and extension. All these features are present in the studied taxa.
In summary and following previous authors, Armella et al. infer a plantigrade foot posture for the specimens reported. Concerning Colbertia lumbrerense, Armella et al.'s results are different from those proposed by Mariano Bond in 1981, who suggested a semidigitigrade-digitigrade posture based on the presence of long and slender tibia and fibula, and a long tuber calcis observed on PVL 6218 (poorly preserved). At this point, the new tarsal remains (PVL 6227) allowed Armella et al. to consider structures and features more directly related to the foot posture, many of which support a plantigrade condition. This is in agreement with the posture inferred for Colbertia magellanica.
Bruce Shockey and John Flynn inferred that Notoungulates evolved from fully plantigrade and pentadactyl (common in the Palaeocene and Eocene) to digitigrade forms (frequent in Oligocene and younger faunas). Furthermore, the foot posture was found to be related with the degree of hypsodonty along the Cenozoic, with high percentages of plantigrade and brachydont taxa in the Eocene, whereas digitigrade morphologies and hypsodont taxa are more common from the middle–late Cainozoic across much of South America. The specimens from Lower and Upper Lumbrera Formation show morphologies consistent with this scenario, supporting the previously proposed trend.
The proximal tarsals of Notoungulates from the Lower and Upper Lumbrera formations are suitable materials for detailed taxonomic analyses. Several morphological features can be clearly distinguished in most cases and provide new anatomical data for several Eocene Typotherian Notoungulates.
The comparison of the tarsals of Colbertia lumbrerense with Colbertia magellanica reveals several differences, mainly on the astragalus. Most of them include variations on the development and arrangement of articular facets, and the small size of the dorsal astragalar foramen in the specimen of the Lower Lumbrera Formation. These differences between the two species of the genus Colbertia highlight the importance of proximal tarsals as a source of useful diagnostic traits to further characterise species.
The PVL 4292, referred to as Typotheria indet., shows morphological affinities with basal interatheriid taxa. Notwithstanding these similarities, the notable larger size of PVL 4292 compared to the overall small body sizes of Eocene Interatheriids and the absence of that family so far in the highly fossiliferous levels of the Lower and Upper Lumbrera formations preclude an indisputable taxonomic assignment.
Concerning 'Campanorco inauguralis', the tarsals exhibit a combination of unique features useful for diagnostic purposes. Armella et al.'s observations indicate that there is no particular morphological evidence for a close phylogenetic relationship with the Mesotheriidae clade, although it should be noted that the considered features were not included in a cladistic analysis so far. Moreover, 'Campanorco inauguralis' shows a 'reversed alternating tarsus' condition, also observed in Leontiniidae, 'Notohippidae', Toxodontidae, and some Typotherians. However, the spectrum of singularities exhibited by these forms precludes, at this point, the assessment of its relationships in the context of the Palaeogene radiation of Typotheria. In this sense, it is necessary to expand the comparative basis among Eocene Notoungulates, so that their evolutionary significance can be fully appreciated.
On the other hand, the detailed anatomical description in a morphofunctional context performed here allows Armella et al. to infer a plantigrade foot posture for the studied specimens. These inferences could potentially be used as functional proxies for palaeoecological purposes, especially informative for the study of the early radiation of these Notoungulate faunas.
See also...






Online courses in Palaeontology.
Follow Sciency Thoughts on Facebook.
Follow Sciency Thoughts on Twitter.