
The origin of Euarthropoda, including the extant groups Myriapoda, Insecta, Eucrustacea, and Chelicerata and different fossil representatives, lies more than half a billion years ago. In this long time, an enormous species richness and morphological diversity evolved in the different ingroups. Within Euarthropoda, the lineage of Chelicerata is often somehow treated as the more ‘basal’ or ‘primitive’ side of the tree and is thought to be more ancestral, especially in comparison to Mandibulata, the other major lineage within Euarthropoda. This view is most extremely applied to Xiphosurida (Horseshoe ‘Crabs’) but also to now extinct groups such as Eurypterida (Sea Scorpions) or the Spider-like group of Trigonotarbida. Horseshoe ‘Crabs’ (though the old fashioned ‘Sword Tails’ would be less ambiguous) have often been treated as ‘living fossils’ (a very unscientific term). They have also been assumed to be a kind of proxy for the early terrestrialisation within Euchelicerata. Yet, this interpretation is most likely incorrect. It is unlikely that the stem species of Euchelicerata was already amphibious, hence the terrestrial behaviour of modern representatives of Xiphosurida has most probably evolved independently. It needs to be emphasised that modern representatives of Xiphosurida are not direct proxies for the ancestor of Euchelicerata, but possess their own specialisations (as all living groups do).
One reason why the lineage of Chelicerata is assumed to be primitive might be the organisation of the feeding apparatus in most representatives. In different textbooks, the impression is given that these forms lack ‘proper’ mouthparts, while ingroups of Mandibulata have a ‘full’ set of mouthparts. One important German text book states that representatives of Chelicerata bear structures for acting in feeding but would lack true antagonistic jaws. He also states that mostly only the second pair of appendages and rarely the third and fourth one is incorporated in the feeding apparatus in most forms. This statement most likely refers to the state in Scorpions and other arachnids, but appears to ignore the state in Xiphosurida where appendage pairs 1–7 are involved in the feeding apparatus. In other cases the description of the feeding apparatuses appears to refer to only certain ingroups. For example the statement that there are always three pairs of mouthparts in Mandibulata is clearly correct for Insecta, but ignores maxillipeds in Chilopoda or Decapoda, or complex thoracic feeding apparatuses as for example in Branchiopoda (Fairy Shrimps). Hence, such textbook statements are at best oversimplified and tend to be based more on general assumptions than on direct observations.
In a paper published in the journal PeerJ on 13 August 2020, Carolin Haug of the Department of Biology and GeoBio-Center at Ludwig-Maximilians-Universität München, provides functional morphological interpretations of the feeding apparatuses (i.e. all external structures involved in feeding) of two supposedly ‘primitive’ Chelicerate groups, namely of Eurypterida and of Trigonotarbida and present them in the context of evolution in Euchelicerata. With this, she aims at providing a framework to evaluate the presumed ‘primitiveness’ of the feeding apparatuses of modern representatives of Euchelicerata.
For this study, fossil material from different museum collections was investigated, including the invertebrate palaeontology collection of the Yale Peabody Museum of Natural History, New Haven, the Natural History Museum, London, the Naturhistoriska riksmuseet, Stockholm, the Harvard Museum of Comparative Zoology, Cambridge, Massachusetts, and the Royal Ontario Museum, Toronto. Every specimen of Eurypterida and Trigonotarbida in these collections was briefly inspected, and those preserving morphological details of interest for this study were documented.
The specimens of Eurypterida in the Yale Peabody Museum of Natural History collections had been collected by a private collector, Samuel Ciurca, from late Silurian deposits in New York State, USA, and southern Ontario, Canada, and donated by him to the Yale Peabody Museum; it is by far the largest collection of Sea Scorpions worldwide. The Sea Scorpions in the Natural History Museum collections originate from different late Silurian deposits: few specimens come from New York State and from Scotland. Also one Sea Scorpion from the Harvard Museum of Comparative Zoology collections comes from the Silurian of Scotland. Most specimens in the Natural History Museum collections come from Estonia, more precisely from the village Rootsiküla on the island of Saaremaa (also called Ösel). From the same Estonian locality, also many specimens of Eurypterida in the Naturhistoriska riksmuseet collections and most in the Harvard Museum of Comparative Zoology collections originate, as well as from the island of Gotland, Sweden. The specimens from Saaremaa/Ösel and Gotland are insofar exceptional as of most of these the surrounding limestone matrix had been dissolved already in the 19th century by Gerhard Holm, resulting in isolated specimens with only their cuticle being preserved. Subsequently, the specimens had either been mounted between two large cover slips, or between a microscopic slide and a cover slip (dry or with Canada balsam), or they have been fully embedded in resin. All these methods allow to access dorsal as well as ventral structures.
The specimens of Trigonotarbida in the Royal Ontario Museum collections come from the Upper Carboniferous Mazon Creek, Carbondale Formation, IL, USA. The Natural History Museum collections houses specimens of Trigonotarbida preserved in Rhynie Chert, Lower Devonian of Scotland, the rest of the material stems from the British Coal Measures, Upper Carboniferous.
Comparative material of extant species came from the former teaching collection of the University of Ulm (now at the University of Rostock) and the teaching collection of Ludwig-Maximilians-Universität München.
The appendages of seven segments clearly contribute to the feeding apparatus in Eurypterida: chelicerae, five subsequent pairs of legs (often called walking legs), and the plate-like appendage pair of the seventh post-ocular segment, the metastoma. The appendages are arranged in a circle around the central area of the feeding apparatus, with their median parts being very close together. The basipods (coxae in chelicerate terminology) of the appendages of post-ocular segment 6 (the leg pair right in front of the metastoma) are much larger than those of the other appendages, forming roughly a rectangle, but additionally with a pronounced endite. The metastoma covers part of these basipods as well as the median gap between the left and right appendage.

Post-ocular appendages 2–6, that is all post-cheliceral legs, bear a more or less pronounced armature on the endites protruding from the median edges of their basipods. The armature differs between the appendages. The spines on some of the basipods (probably of the further anterior ones, but there appear to be species-specific differences) are thinner than on others. The basipods of post-ocular appendage 6 bear the stoutest spines, the entire basipodal median edge appearing strongly sclerotised, recognisable from its very dark colour.

Also the armature on the same basipod is differentiated (also here with species–specific differences). The spines closer to the anterior edge of the basipod (in relation to the orientation of the body of the animal) are stouter and partly also longer, while those closer to the posterior edge of the basipod are thinner and smaller. This differentiation can be very pronounced, with very stout and large teeth, elongate and pointed spines, and thin setae on the same basipod. The armature on the median edge of the basipods can occur in two (or possibly three?) rows. The remaining surface of the basipods can be covered with setae of other surface structures.

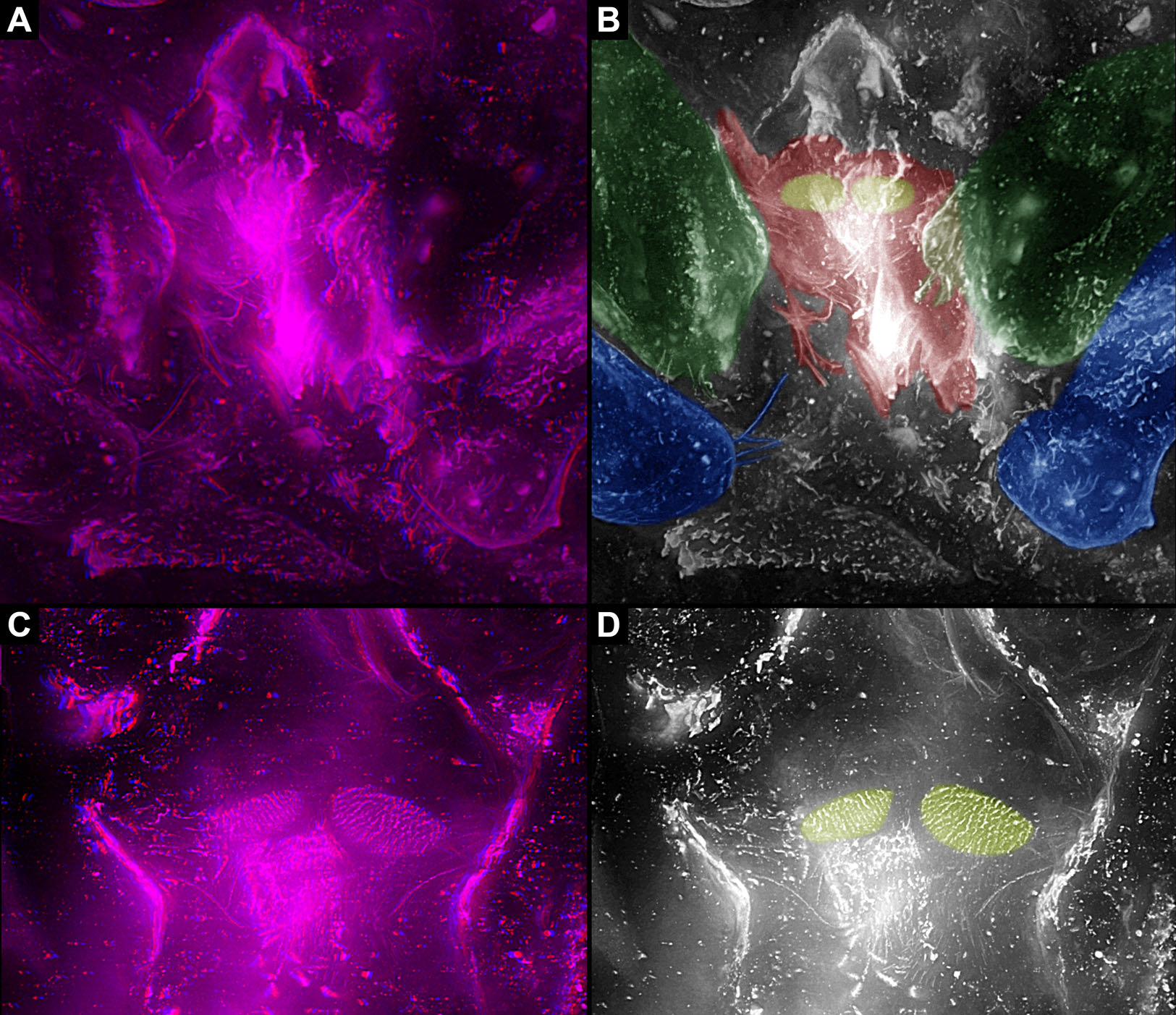
In Trigonotarbida, the appendages of three segments contribute to the feeding apparatus: chelicerae, pedipalps and first pair of walking legs. The proximal parts of these appendages sit close together. The pedipalps are proximally armed with a row of short spines. The basipods of the first pair of walking appendages bear a prominent endite medially, which again bear setae. The endites are in some fossils positioned very closely together, in others further apart, which may point to a high movability during life. Additionally, there are two distinct oval fields of densely arranged short setae next to each other in the area between chelicerae and pedipalps. Another less distinct field of similar setae appears to be positioned slightly posteriorly to the two oval fields. Based on the three-dimensional position information, these fields are probably situated on the hypostome (though often termed ‘labrum’).

While Haug's study focused on the feeding apparatus of different groups of Euchelicerata, a proper character polarisation demanded an outgroup comparison. For this purpose it wss especially important to include now extinct groups as those may exhibit character states no longer present in the extant fauna.

The supposed sister group to Euchelicerata is Pycnogonida (Sea Spider), these together forming the Chelicerata. While being quite speciose today, the general body organisation is the same in all extant species of Pycnogonida, with a highly reduced posterior body area (the terms pro- and opisthosoma cannot be applied) and a strongly modified feeding apparatus adapted to suctionfeeding. The fossil record of the Pycnogonida dates back to the Cambrian, so more than half a billion years ago. However, as already at this time the morphology appears derived (and partly also as the Cambrian representatives of Pycnogonida are exclusively larvae), it does not provide relevant information about the evolution of the feeding apparatus in the lineage towards Euchelicerata. It is necessary to take a look at the feeding apparatus in supposed early representatives of Megacheira, the so-called ‘short great-appendage Arthropods,’ among which the sister group to Chelicerata is assumed by some authors.

Recent reinvestigations of different of these species as well as older publications provide a sound basis for the reconstruction of the feeding apparatus in the ground pattern of Megacheira. At this evolutionary stage, the entire set of appendages was included into the feeding apparatus (possibly including the hypostome). While the first pair of appendages was specialised for grasping the prey, the second to last pair of appendages all exhibit the same general morphology, serving for swimming and feeding at the same time. While slight modifications in the size of the second appendage pair of Leanchoilia superlata might indicate a starting specialisation to a mouthpart, the ‘whole-body-feeding’ method represents the original condition in Megacheira and presumably in Chelicerata.

This also holds true if assuming that short great-appendage Arthropods branched off earlier along the evolutionary lineage. The polarisation of characters remains the same. Also several other early representatives of Euarthropoda possess similar characters in their feeding apparatuses, most prominently opposing basipods with strong spines, but are not discussed further by Haug.

The feeding apparatus of Pycnogonida is adapted to suction feeding and highly derived already in the fossil forms. This specialised feeding apparatus is an autapomorphy of Pycnogonida.
In Euchelicerata the systematic affinities are still in a certain state of flux and the feeding apparatus cannot be properly reconstructed for all early fossil representatives. The feeding apparatus and in general the tagmosis pattern can be reconstructed to a certain extent for the species Offacolus kingi, Dibasterium durgae, and Weinbergina opitzi, successively splitting off the evolutionary lineage towards the remaining representatives of Euchelicerata.

At the level of Euchelicerata, a characteristic division into an anterior and a posterior tagma usually termed prosoma and opisthosoma appears for the first time, supposedly as an autapomorphy for this group. The subdivision into these two distinct tagmata is characterised by dorsal and ventral structural changes.
Dorsally, the segments of the anterior tagma form a uniform shield without any visible segmental borders. The segments of the posterior tagma possess separate tergites in the ground pattern of Euchelicerata. Ventrally, the tagmatisation is characterised by a ‘division of labour’ between the different appendages. While in short great-appendage Arthropods all appendages were still involved in feeding, in the ground pattern of Euchelicerata the feeding apparatus becomes shorter; only the appendages of the anterior tagma contribute to the feeding apparatus, in addition to their locomotory (walking) function. The more posterior appendages serve for swimming; if they also possessed structures for oxygen exchange remains currently unclear.

The appendage of post-ocular segment 7 (the pre-genital segment) differs morphologically in Offacolus kingi, Dibasterium durgae, and Weinbergina opitzi, being significantly smaller in the first two species. Yet, as Weinbergina opitzi possesses an appendage on this segment similar to the preceding ones with apparently locomotory function, this was presumably the ground pattern condition for Euchelicerata. Hence, the feeding apparatus in the ground pattern of Euchelicerata most likely consists of (possibly the hypostome and) chelicerae and six pairs of walking appendages.

Neochelicerata is an ingroup of Euchelicerata, the ‘crown group’ (most inclusive group with extant representatives). It has been characterised as certain aspects of body organisation have not evolved in Euchelicerata yet, but are present in the ground pattern of Neochelicerata. The ground pattern condition of Neochelicerata is mainly reconstructed based on the morphology of Xiphosurida.
The body organisation in general and the feeding apparatus in particular are very similar in the ground pattern of Neochelicerata to that of Euchelicerata. Also here the anterior tagma dorsally bears a uniform shield. The tergites of the posterior tagma are fused into an entire dorsal shield, the thoracetron, in Xiphosurida. However, this condition appears to be an autapomorphy of Xiphosurida, while in the ground pattern of Neochelicerata the tergites were most probably still separate.
Also ventrally, the ground pattern condition of Neochelicerata is largely the same as that of Euchelicerata. The appendages of the anterior tagma, (possibly the hypostome and) chelicerae and the following six pairs of appendages, are incorporated into the feeding apparatus. However, the last of these appendage pairs, the chilaria (appendages of the pre-genital segment or post-ocular segment 7), consists of only the shovel-shaped most proximal part (basipod in neutral euarthropodan terminology, usually called coxa in Chelicerate terminology); the walking part (endopod) is lacking. With this, the appendages of post-ocular segment 7 no longer perform a combined feeding-and-walking function, but instead close the feeding apparatus from its posterior end to reduce the loss of food in Xiphosurida. This condition is possibly already present earlier as it looks very similar in a species splitting off the evolutionary lineage before the node of Neochelicerata, Venustulus waukashensis. Therefore, in the ground pattern of Neochelicerata (or slightly earlier) the feeding apparatus became further specialised, consisting of (possibly the hypostome,) chelicerae, five pairs of walking appendages and chilaria.
The exact relationships of Eurypterida, Arachnida, Xiphosurida, and other exclusively fossil groups such as Chasmataspidida are still not entirely The feeding apparatus of Eurypterida (and possibly already of Metastomata if this is a natural group) shows a stronger specialisation than that in the ground pattern of Neochelicerata, but is still composed of (possibly the hypostome and) the appendages of the same segments (chelicerae, following five pairs of appendages, metastoma). The further posterior segments are also here not involved in feeding or locomotion; the corresponding appendages most probably became (partly) internalised and fulfilled respiratory function.

Also in Sea Scorpions, feeding and locomotion appears to be performed by the same appendages. However, Paul Seldon, who described the functional morphology of Eurypterus tetragonophthalmus in great detail, assumed that only the posterior appendages were used for locomotion to avoid coordination problems between the differently long appendages (though this assumption does not have to account for all species of Eurypterida due to their morphological differences). He also assumed that there is a task differentiation in food handling between the anterior and posterior legs, the anterior ones gathering food, while especially the last pair crushed hard food particles. This differentiation is corroborated by the specimens investigated in this study, as the basipods of the different appendages are equipped with different types of teeth on their median edges, best visible when (almost) the entire feeding apparatus is preserved in situ. In some specimens the teeth on the basipods of the further anterior appendages are thinner and appear less robust than those on the basipods of the fifth (last) pair of walking legs (appendage pair of post-ocular segment 6). The antero-median edges of the latter are equipped with strong teeth and reach under the metastoma. They are also significantly elongated in anterior-posterior axis, probably to achieve a larger biting force. With this, sea scorpions possessed fully functional antagonistic jaws comparable to the condition in many Crustaceans, but convergently evolved.
In addition to the differentiation of armature on the basipods of different appendages, on the same basipod the armature is also differentiated. The teeth further anterior on the median edge of the basipod appear stronger than the further posterior ones on the same basipod. Paul seldon described this differentiation for Eurypterus tetragonophthalmus, and discussed that some of the teeth or spines would have been movable while others were not. The presence of movable teeth may be species specific based on the observations in this study, but the general pattern of differentiated armature on the same basipod apparently occurs in different species of Eurypterida. This differentiation appears very similar to the situation in Mandibulata, in which the mandibles bear the pars incisivus and the pars molaris, that is two regions with rather different armature. Apparently, also this specialisation evolved convergently.
The metastoma (appendage of pre-genital segment or post-ocular segment 7) in representatives of Eurypterida basically fulfils the same function as the chilaria in representatives of Xiphosurida, it closes the feeding apparatus from posteriorly. Yet, in Sea Scorpions this is only a single plate (but which can have various shapes), so most probably the conjoined basipods of formerly free appendages. Some species show a notch in the anterior area of the metastoma, which may represent a remnant of the not completed fusion process. Unfortunately, no extremely early ontogenetic stages are preserved (late embryos to hatchlings, at least not well enough to allow investigation of the development of the metastoma.
In addition to the closing of the feeding apparatus, the metastoma appears to provide a kind of guide rail for the movements of the basipods of the appendage pair right in front of it. This function is comparable to that of the paragnaths in Eucrustaceans, which are elevations of the sternite of the mandible segment and guide the movement of the mandibles/ Again, this morphological similarity evolved most likely convergently.
The condition of the metastoma as a single plate and its covering of the proximal area of the posterior appendages leads to a more tightly closed feeding apparatus in comparison to that in Xiphosurida and the ground pattern condition of Euchelicerata. Considering that representatives of Eurypterida may have had an amphibious lifestyle, a more closed feeding apparatus could have been a predisposition for going on land, as food may get lost more easily on land than in the water, where it sinks slower when it is not grabbed tightly enough. The specialised feeding apparatus of Sea Scorpions in its highly differentiated morphology is probably best interpreted as an autapomorphy of Eurypterida or even of an ingroup.
In the following, Scorpions are taken as a first example for Arachnida. Scorpions have been assumed to be the sister group to the remaining groups of Arachnida. Yet, in recent studies, Scorpions resulted as sister group to Megoperculata; in these analyses Sea Sorpions were not included (which was mostly not possible due to the type of analysed characters). This deep ingroup position of Scorpions may be an artefact resulting from the lack of proper character polarisation due to the absence of Eurypterida, but this problem cannot be further discussed here.
In Arachnida, the posterior border of the feeding apparatus lies further anteriorly than in the previously discussed groups. In modern Scorpions (Scorpiones), only the first four appendage-bearing segments (and possibly the hypostome) are involved in the feeding apparatus: chelicerae, pedipalps, and two pairs of walking appendages. The basipods of these two pairs of walking appendages are antero-medially elongated into a pronounced endite, which reaches far anteriorly, closing the feeding apparatus from the posterior end. The median areas of the basipods of walking appendage pairs 3 and 4 are oriented almost anteriorly, but apparently not included into the feeding apparatus. Also the sternum, most likely the embryonically fused appendages of the seventh post-ocular (pre-genital) segment, is oriented anteriorly, but not involved in the feeding apparatus. Dorsally, the shield extends further posteriorly, including the segments bearing chelicerae, pedipalps, all walking appendages and possibly also the sternum-bearing segment. Hence, the posterior border of the feeding apparatus in Scorpiones no longer corresponds to the posterior border of the dorsal shield, in contrast to the presumed ground pattern condition in Euchelicerata, Neochelicerata, and Metastomata.
However, the condition of the feeding apparatus in modern Scorpions differs from that in early fossil Scorpions. In early Scorpions, the feeding apparatus extends posteriorly only to the pedipalps. The walking appendages do not bear enditic protrusions, hence do not appear to have been involved in the feeding process. If this condition is the ground pattern condition of Scorpionida, the group including Scorpiones and different fossil Scorpions, the feeding apparatus in modern Scorpions would be an autapomorphy of Scorpiones.
In Trigonotarbida, the Chelicerae, pedipalps and first pair of walking appendages contribute to the feeding apparatus. The basipods of the pedipalps and especially of the first pair of walking appendages possess endites medially, which were probably used for food manipulation. This condition is unknown from other Arachnids.
The phylogenetic position of Trigonotarbida is still unclear. In different analyses, they have already resolved, for example, as sister group to Ricinulei or as closely related to Megoperculata. Similarities to the one respectively the other group occur, for example, in the filtering structures on the mouthpart, details on the pedipalps, or the general body organisation.
The feeding apparatus does not provide clear arguments for placing Trigonotarbida near the one or the other group of Arachnida. In most representatives of Arachnida, the feeding apparatus only consists of (possibly a hypostome,) chelicerae and pedipalps. In some of them, the basipods of the pedipalps are more or less closely connected medially. The basipods of the pedipalps are in many Web Spiders involved in the feeding process, using median projections for mastication (sometimes referred to as gnathocoxae or maxillae). In certain cases, a lower lip contributes to the feeding apparatus from posteriorly. In some groups, all these structures form a single, tightly connected unit. If this short condition of the feeding apparatus would be present in all representatives of Arachnida besides Trigonotarbida, two options for the ground pattern of Arachnida would be possible: either the feeding apparatus of Trigonotarbida would represent the ground pattern condition of Arachnida, or the very short condition of the other representatives of Arachnida would be the ground pattern condition and Trigonotarbida autapomorphically elongated the feeding apparatus.
However, the entire situation is more complicated due to the feeding apparatus of Opiliones (Harvestmen). In Harvestmen, the feeding apparatus includes also the first pair of walking legs, which bears endites on the basipods. Also the second pair of walking legs bears endites, at least in certain Harvestmen, which may have a supporting function in the feeding process. The presence of a preoral chamber, stomotheca, formed by all these endites in Opiliones and Scorpionida has led to the suggestion a sister group relationship between these two groups (together forming Stomothecata). The endites of the first pair of walking appendages in Harvestmen look rather similar to those in Trigonotarbida, and in general also the composition of the feeding apparatus would be similar between the two groups/However, the morphology in Harvestmen is highly variable, and it is unclear how the feeding apparatus in the ground pattern of Opiliones looks. This together with the still unresolved phylogenetic position of Opiliones does not allow to make a reliable assumption about the feeding apparatus in the ground pattern of Arachnida.
The feeding apparatus in different groups of Euchelicerata is far from primitive, but is in fact a highly specialised system in each group. During evolution, the feeding apparatus became progressively shorter in Euchelicerata, though the evolution within Arachnida remains still unclear due to unresolved phylogenetic relationships. The shortness of the feeding apparatus is not an ancestral character, but in fact a highly derived one, probably evolved in adaptation to new requirements resulting from habitat changes such as terrestrialisation. Sea scorpions possess true antagonistic mouthparts with differentiated armature and a guide rail system. These characters are all similar to the condition in the mandibles of Mandibulata, apparently as result of convergent evolution. Representatives of Trigonotarbida show similar specialisations concerning their feeding apparatuses to Harvestmen.In conclusion, the supposedly ‘primitive’ groups Eurypterida and Trigonotarbida are astonishingly specialised.
See also...






Online courses in Palaeontology.
Follow Sciency Thoughts on Facebook.