
The Cheiracanthidae are a family of Acanthodiform Acanthodians that considered to be represented by two genera based on Articulated Fish, Cheiracanthus and Homalacanthus; another genus, Markacanthus, is known only from isolated scales. Cheiracanthus spp. were abundant in Middle Devonian Vertebrate faunal assemblages of northern Europe, with isolated scales described from the Baltic countries and Russia, and articulated Fish from Scotland and Spitsbergen. Only one valid new species based on articulated specimens, Cheiracanthus peachi, from Scotland, has been described since the nineteenth century. Rare scales have been recorded from Greenland which are now recognised as Cheiracanthus intricatus, but records of Cheiracanthus from elsewhere in the world are now considered incorrect or dubious: scales from Antarctica described in 1968 were subsequently assigned to the Diplacanthid Milesacanthus antarctica, and Cheiracanthus? costellatus, from the Emsian of eastern Canada is most likely a Diplacanthiform. The only other Cheiracanthid genus is Homalacanthus, with the type and only valid species being Homalacanthus concinnus from the Frasnian of Quebec. Protogonacanthus and Carycinacanthus were also assigned to the Cheiracanthidae, but are considered to be Acanthodids, based on their unornamented scale crowns; also, Carycinacanthus is now considered to be a junior synonym of Acanthodes. It has been suggested that the Cheiracanthids are a grade within the Family Acanthodidae, but are currently regarded as being a Family within the order Acanthodiformes based on their endoskeletal structure and scale ornament.
In a paper published in the journal Palaeontologia Electronica on 10 April 2020, Carole Burrow of Geosciences at the Queensland Museum, Jan den Blaauwen of the University of Amsterdam, and Michael Newman of Haverfordwest in Pembrokeshire, Wales, redescribe three long-known species of Cheiracanthus from the Middle Devonian of Scotland.
Cheiracanthus murchisoni was one of the first Devonian Fish to be described. It was erected by Louis Agassiz, in 1835, based on specimen IGUN.66 in the Neuchâtel Museum collection (now MHNN FOS39) from Gamrie in Aberdeenshire. Cheiracanthus minor was erected in the same work, figuring one specimen. This specimen was part of the Traill collection and is now considered lost. Agassiz later erected Cheiracanthus microlepidotus based on three specimens from Lethen Bar in the Highlands, NHMUK P.544 and MHNN FOS 40 and 41. Cheiracanthus microlepidotus is now considered to be a junior synonym of Cheiracanthus murchisoni (i.e. a second name given to the same species in error). Cheiracanthus minor was also considered to be a junior synonym of Cheiracanthus murchisoni by later Victorian palaeontologists, though as the original material has been lost this is hard for modern workers to assess.

Type material of Cheiracanthus murchisoni, and its junior synonym Cheiracanthus microlepidotus. (1)-(3) Holotype MHNN FOS 39 from Gamrie: (1) specimen in present condition; (2) closeup of midbody scales; (3) image in Agassiz (1833-43). (4)-(9): Syntypes of the junior synonym Cheiracanthus microlepidotus from Lethen Bar: (4), (5) NHMUK PV P544; (6), (7) MHNN FOS 40; (8), (9) MHNN FOS 41. Scale bars equal 20 mm. Burrow et al. (2020).
Cheiracanthus grandispinus was erected by Frederick M’Coy McCoy in 1848 but not figured at that time. In 1855, he figured the holotype specimen SM H4423 and gave a fuller description, including detail of the ornamentation of the scale crowns. Both Philip Egerton in 1860 and Ramsay Traquair in 1888 concluded that Cheiracanthus grandispinus was a valid species. No work of note has since been done on the species.

Cheiracanthus grandispinus holotype SM H4423 from Orkney. (1) mirrored illustration of specimen and scales from M’Coy (1855); (2) specimen in present condition. Scale bar equals 20 mm. Burrow et al. (2020).
Cheiracanthus latus was first erected and figured by Philip Egerton in 1861, with his description based on some 50 specimens from Tynet Burn in Moray. He figured only one specimen, but that half of the original nodule is now lost. However, the other half (NHMUK PV P3253) has been located and is now defined as the holotype.

Cheiracanthus latus, 1861 holotype from Tynet Burn. (1) NHMUK PV P3253, half of specimen not previously figured; (2) half of the holotype specimen illustrated by Egerton (1861), now lost. Scale bar equals 20 mm. Burrow et al. (2020).
In Orkney, Cheiracanthus first appears in the Sandwick Fish Bed Member. This horizon is dominated by the species Cheiracanthus murchisoni, with Cheiracanthus grandispinus being much rarer. Both species continue into the overlying Upper Stromness Flagstone Formation, with Cheiracanthus grandispinus surviving the longest, into the middle of this formation. Cheiracanthus latus is unknown in Orkney. Another species, Cheiracanthus peachi, is found much higher, in the upper part of the Rousay Flagstone Formation.

(1) Site map indicating the most important localities in northern Scotland where Middle Devonian Cheiracanthus specimens have been collected, land in white, current shoreline in blue, alluvial plains in yellow, alluvial fans in ochre; main lake area in blue; (2) biostratigraphic table of Cheiracanthus species occurrences in the Orcadian Basin of northern Scotland. Burrow et al. (2020).
In Caithness, Cheiracanthus grandispinus appears first and is confined to the Achanarras Fish Bed Member. Cheiracanthus murchisoni and Cheiracanthus latus are unknown in Caithness. As in Orkney, Cheiracanthus peachi is found up high in the stratigraphical sequence, in the upper part of the Mey Flagstone Formation. At least one undescribed Cheiracanthus species occurs below the Achanarras Fish Bed Member in the Lybster Flagstone Formation.
In the nodule beds of the Moray Firth, stratigraphically equivalent to the Achanarras Fish Bed Member, Cheiracanthus murchisoni, Cheiracanthus grandispinus, and Cheiracanthus latus are present, except for Edderton where Cheiracanthus latus is absent. Some of the nodule beds at Cromarty might be a little lower than the Achanarras Fish Bed Member and possibly contain the same Cheiracanthus species that is present in the Lybster Flagstone Formation. Acanthodians are unknown from higher strata in the Moray Firth area.
The family Cheiracanthidae, as currently defined, comprises Acanthodiforms with a short branchial region completely covered by slender branchiostegal rays; jaw cartilages formed of a single mineralisation composed of calcified cartilage blocks; palatoquadrate with median fenestra; mandibular splints absent; 4-6 sclerotic bones; multicuspid denticles/gill rakers on some of the branchial arches; high slender scapular shaft; ossified coracoid and procoracoid; in spines with smooth rounded leading edge ridge separated by groove from smooth or ridged lateral surfaces; dorsal spine with mineralised basal cartilage; no prepelvic or admedian fin spines; polygonal tesserae on cranial roof; ornamented scale crowns.
Cheiracanthus murchisoni is a species of Cheiracanthus, with more than 20 contiguous long, robust, branchiostegal rays with straight posterior ends; several fine thin rays above the main branchial cover; fine thin gular rays below the jaws; slender, dorsally-tapering scapular shaft, height 10 times its minimum width; spiracle with pair of comma shaped valves; pectoral spine with low sharp-crested ridges along lateral faces; scale crowns ornamented with non-branching subparallel ridges extending from the anterior edge and terminating before mid-crown; some scales with a posterior median pit on the crown; scale crown growth zones with straight posterolateral edges.
The dorsal fin spine is positioned halfway between the pelvic and anal spines. On all fish, the pelvic spines are always the shortest, about 60 percent of the length of the dorsal spine. The anal spine is intermediate in length between that of the dorsal and pelvics, about 70 to 75 percent the length of the dorsal spine. The pectoral spine is variable in length, and can be shorter than, equal in length, or longer than the dorsal spine.
The lower jaw Meckel’s cartilages are always mineralised as a single unit as are the palatoquadrates, but these are less frequently mineralised, and are also often not preserved intact but as scattered patches. Burrow et al. note that the fossa for insertion of the adductor muscle on the lateral side of the Meckel’s cartilage extends about half the length of the jaw. The ventral margin of the cartilage is thickened to form a ridge along this edge. The small preglenoid process is quite rounded, and the articular cotylus is a shallow embayment, slightly longer than wide. Tooth-like structures are present in the mouth. These are possibly eqivalent to similar elements in Homalacanthus concinnus, where they are distributed in rows near the upper edge of the Meckel’s cartilage and the lower edge of the palatoquadrate.

Cheiracanthus murchisoni head and branchial region morphology. (1), (2) NMS G.1884.60.3 from Tynet Burn. (3) NMS G.2019.9.24 from Cruaday Quarry, Orkney. (4) NRM P1651 from Gamrie, sclerotic ring. (5) NRM P1654 from Tynet Burn, sclerotic ring. (6) NMS G.2000.65.2 from Tynet Burn. (7) NMS G.2019.14.2 from Tynet Burn. (8), (9) NRM P1560 from Gamrie: (8) detail of branchiostegal rays; (9) impression of spiracular valve. Abbreviations: addf, adductor muscle fossa; artc, articular cotylus; br, branchiostegal rays; ch, ceratohyal; gr, gular rays; Mc, Meckel’s cartilage; pdbr, posterodorsal branchiostegal rays; pq, palatoquadrate; pqv, palatoquadrate vacuity; pregl, preglenoid process; scl, sclerotic plate; t-l, tooth-like elements. Arrows indicate anterior. Scale bars equal 10 mm in (1), (3), (6), (7); 5 mm in (4), (5); 2 mm in (8), (9). Burrow et al. (2020).
The two main dermal elements of the head and branchial region are the orbital bones and the branchiostegal rays. The former comprise 4-6 sclerotic plates, not circumorbital bones. Disarticulated sclerotic plates show that they are short, curved, and mesially concave. In articulation they are close-set. The internal surface is smooth, but the external surface on at least some specimens was ornamented with fine, sinuously radiating ridges. The ornament layer appears very thin and is rarely preserved intact.
There are about eight dermal branchiostegal rays below the angle of the jaw and 16 or more above the jaw angle; some of the larger rays have an ornamentation of two ridges running parallel to their long edges. The most dorsal branchiostegal rays of the main cover are fairly flat with no abrupt bends to the ventral on the posterior ends of the individual rays. Burrow et al. observe that there are also long thin rays above the main branchial cover. These are much more fragile than the branchiostegal rays and are rarely preserved. Short delicate rays are present between the dorsal area of main branchial cover and the shoulder girdle. Based on their position and circular arrangement, Burrow et al. interpret two small sickleshaped bones above the main branchial cover on NHMUK PVP.43273 as the dermal spiracular valve. The internal pseudobranch is also seen as an impression on NRM P1650 and in thin section on NMS G.2019.3.6.15. Thin gular rays are also present below the jaws. Elements not previously identified are denticles in the branchial region, presumed to have been borne on the branchial arches.
Thin sections show that the jaws and at least some other endoskeletal structures in these regions are preserved as a layer of calcified cartilage, often formed of contiguous blocks which are sub-rectangular in vertical section. In other areas, the cartilage layer is formed of separated irregularly shaped blocks. Rings and lines of Liesegang are sometimes visible within the calcified cartilage blocks. The jaw cartilages are preserved as a single-layer of calcified cartilage blocks enclosing a calcitic core, presumed to have originally been unmineralised cartilage. Both dorsal and ventral edges of the jaws are thickened, but still encased with just a single layer of calcified cartilage blocks.

Cheiracanthus murchisoni head and branchial region histology. Thin sections of NMS G.2019.3.6. (1) NMS G.2019.3.6.2, Meckel’s cartilage; (2) NMS G.2019.3.6.7, calcified cartilage block showing rings of Liesegang; (3) NMS G.2019.3.6.3, separated calcified cartilage blocks overlain by dermal tessera; (4) NMS G.2019.3.6.4, calcified cartilage and ?subcylindrical sensory line scale; (5) calcified cartilage of palatoquadrate with dermal tesserae and pair of scales in vertical longitudinal section, edging sensory line; (6) NMS G.2019.3.6.6, ridge on dorsal edge of palatoquadrate cartilage, calcite infilling; (7) NMS G.2019.3.6.9, pair of sensory line scales, oblique section through base and lower crown; (8) NMS G.2019.3.6.12, dermal tesserae, vertical transverse sections; (9) NMS G.2019.3.6.9, pair of sensory line scales, vertical transverse section; (10) NMS G.2019.3.6.7, ceratohyal and another branchial arch; (11) NMS G.2019.3.6.6, branchial arches; (12), (13) NMS G.2019.3.6.9, ?tesserae and ?denticle of the branchial region; (14), (15) NMS G.2019.3.6.5, branchiostegal plates and calcified cartilage, with closeup of branchiostegal transverse section; (15) tessera with polyodontode crown on globular calcified cartilage; (16) NMS G.2019.3.6.4, branchiostegal plates and gular rays; (17) NMS G.2019.3.6.15, spiracular valve. Scale bars equal 1.0 mm. ba, branchial arches; br, branchiostegal plate/ray; cc, calcified cartilage; ch, ceratohyal; d, denticle; gr, gular rays; t, tessera; ?, indeterminate element, possibly subcylindrical sensory line scale. Burrow et al. (2020).
Flat or slightly concave based dermal tesserae are borne directly on the cartilage blocks. The sensory lines on the head are enclosed by paired specialised scales with thin flat bases and thin crowns that arch over and meet along the median line between the two rows of scales. The scale crowns are formed of superposed growth zones. Tesserae above the jaws have a thick base penetrated by Sharpey’s fibres and a dentinous crown showing apposed growth zones.
The anteriormost branchial arches are formed of a two-layered, consolidated calcified cartilage, with the outer layer appearing denser than the inner layer. This two-layered structure perhaps equates with a mesodermal and neural crest derived origin for the outer and inner layers respectively, based on comparison with the development of the gill arches in Skate embryos.
Some of the branchial arches are surrounded by small plates with a long inclined pointed crown, which we identify as branchial denticles. Other larger elements in the branchial region show a vertical section similar to that of the branchial denticles, but have a large pulp cavity and a dentine crown. Burrow et al. identify these elements as equivalent to the supposed gill rakers of the Cheiracanthid Homalacanthus concinnus. Burrow et al. are unable to determine the exact distribution of the elements in Cheiracanthus murchisoni, but refer to them as tooth-like elements as it seems likely that, as in Homalacanthus concinnus, they are only found near the edges of the palatoquadrate and Meckel’s cartilage rather than being associated with the branchial arches. The dermal branchiostegal plates and smaller gular rays are formed of a dense bone; the ornament layer on the branchiostegal plates is formed of superposed dentine layers.

Cheiracanthus murchisoni head and branchial region reconstruction. Burrow et al. (2020).
The scapulocoracoid has a slender tapering scapular shaft with a circular cross section. The minimum diameter of the shaft increases very slightly between the smallest and largest Fish, being 0.9 mm in the 100 mm long NMS G.1965.59.34 and 2.0 mm in the 205 mm long NMS G.1972.23.1. The height of the scapulocoracoid, however, increases more dramatically, being 6.0 mm in the smallest and 19.0 mm in the largest Fish. The ventral edge of the scapulocoracoid is contiguous with the upper surface of the pectoral fin spine. The line of fusion between the scapula and coracoid is convex upwards.

Cheiracanthus murchisoni spines and scapulocoracoid morphology. (1) NMS G.1891.92.307 from Gamrie, specimen showing disposition and relative sizes of fin spines and scapulocoracoid. (2) NMS G.1973.12.120 from Lethen Bar, articulated scapulocoracoid and pectoral fin spine. (3) QMF60005 from Tynet Burn, scapulocoracoid ‘shell’ with calcified cartilage core and calcified separation between molds of scapula and coracoid elements. (4), (5) NMS G.2019.9.27 from Edderton, pectoral spine. (6) NMV P29287 from Gamrie, anal fin spine, web, and basal cartilage. (7) NRM P1654 from Tynet Burn, dorsal fin spine with basal cartilage. (8) NMS G.1972.23.1 from Lethen Bar, dorsal fin spine with basal cartilage. Abbreviations: a.fs, anal fin spine; bc, basal cartilage; cor.m, coracoid mold; d.fs, dorsal fin spine; pect.fs, pectoral fin spine; pelv.fs, pelvic fin spines; sc.m, scapula mold.Arrow indicates anterior. Scale bars equal 20 mm in (1), (2), (6)-(8); 2 mm in (3)-(5). Burrow et al. (2020).
All the fin spines have only one groove on each side, separating the leading edge ridge from the ‘shoulder’ (i.e., rounded upper corner) of the sides. Towards the base of the spine, the groove becomes deep and narrow. The pectoral fin spine has low, sharp-crested lateral ridges, which are sometimes also present on the dorsal fin spines. In all spines the inserted part is short. The pulp or central cavity of the spines is open posteriorly/along the trailing edge for a considerable distance, up to half the total length on the dorsal spine.
The dorsal fin spine s almost straight, laterally compressed, with a broadly rounded leading edge ridge; the sides of the ridge are in line with the shoulders and the flanks. The height of the spine increases relative to the width from proximal to distal. The anal fin spine is slightly recurved and also has a broadly rounded anterior rib; the sides are more rounded than on the dorsal spine. Both dorsal and anal spines sometimes have a mineralised basal cartilage preserved; its presence or absence appears independent of locality.
On the paired pelvic fin spines, the anterior ridge has a more or less triangular shape, and the height to width ratio increases from proximal to distal. The spines are notably asymmetric.
The pectoral spine is strongly curved lengthwise, relative to the other spines. Like the pelvic spines, the spines are slightly asymmetric in cross section, and the anterior ridge has a sharply pointed triangular shape in cross section. The sides are straight, in line with the sides of the anterior ridge.The height:width ratio increases dramatically from proximal to distal, and the spines are more robust than the other spines.
All spines, like those of all Cheiracanthus species, have a thin enameloid outer layer on the leading edge and ‘shoulders’ of the spine, preserved best towards the proximal end, as are the thin sharp-crested lateral ridges. Spines have a wide central cavity, and all spines lack an accessory pulp canal above the central cavity. Osteodentine forms most of the width of the spine proximally. In the leading edge ridge, dentine tubules run perpendicular from the outer surface towards vascular canals running longitudinally. These canals are arranged in concentric, interconnected series, more or less parallel to the outer surface of the spines. Longitudinal and interconnecting canals extend the length and breadth of the whole spine, with smaller canals leading off to open out in the sulcus between the leading edge ridge and the ‘shoulders’ of the spines, as well as into the central cavity. Denteons are formed around the canals in the exserted part, but not in the inserted part; they increase in thickness distally, with the canals subsequently becoming narrower or almost closed. An inner lamellar layer is present proximal to the closure of the central cavity, and increases in thickness distally. A very thin, dense layer lines the central cavity proximally, and separates the osteodentine layer from the inner lamellar layer through the whole spine. Short dentine tubules extend through each of the lamellar layers, perpendicular to the inner surface. Towards the tip of the spine, the central cavity is almost filled by the centripetal growth of the inner layers of dentine.

Cheiracanthus murchisoni NMS G.2019.3.6 from Gamrie, scapula shaft and pectoral spine histology. (1) NMS G.2019.3.6.27, transverse section of scapula shaft. (2)-(6) transmission microscope images of NMS G.2019.3.6.14, 15, 16, 18, 19; (7) drawings of main histological features in thin sections 19-14, from tip to near the base of the exserted part. Abbreviations: co, canal opening; en, enameloid; r, sharp crested lateral ridges. Scale bars equal 0.1 mm in (1)-(3), 0.5 mm in (4)-(6). Burrow et al. (2020).
It has previously been suggested that the scales of Cheiracanthus murchisoni are relatively uniform over the body, but Burrow et al. note some variation. In the anterior half of the body the width of the scales sometimes slightly exceeds the length, whereas posterior to the anal fin spine the length of the scales often considerably exceeds the width, with the maximum length to width ratio about 2:1. On most flank scales, the crown ornament comprises fine subparallel ridges extending back from the anterior margin, which often has a smooth rim on scales of larger Fish. The ridges have a rounded surface, do not branch, and terminate in the anterior half of the crown. The length of the ridges varies depending on the area of the body. with scales above the lateral line tending to have longer ribs than those below. Ridge length also depends on the age of the Fish, with larger/older Fish having relatively longer ridges than smaller/juvenile Fish. Many scales have a median depression longitudinally on the crown surface.

Cheiracanthus murchisoni squamation: light microscope images, all mid-body scales except (9). (1) NMS G.1892.8.1 from Gamrie. (2) NMS G.1891.92.323 from Lethen Bar. (3) NMS G.2019.9.25 from Cromarty, Sutors. (4) NMS G.2019.9.26 from Tarbat. (5) NMS G.1882.16.13 from Cromarty. (6) NMS G.1899.83.5 from Hooveth, Orkney. (7) NMS G.1877.30.2 from Tynet Burn. (8) NMS G.1968.5.3 from Edderton. (9) NMS G.2019.9.27 from Blackpark, tail. (10) NMS G.2019.9.28 from Blackpark, scales with crown pits. Abbreviation: p, pit on posterior scale crown. Scale bars equal 1.0 mm in (1), (2), (7), (8); 0.5 mm in (3)-(6), (9), (10). Burrow et al. (2020).
Rows of scales near the lateral lines and also some in other areas have a median pit in the posterior half of the crown. Some of these pitted scales have a crown that is otherwise identical to the majority of scales, whereas others have a relatively narrow crown with thicker subparallel ribs extending to the posterior of the crown, around a relatively large pit. Scales on the front edge of the tail are larger than body scales and have a totally smooth crown. The neck is concave and smooth all round, with a pair of protuberances posteriorly, just above the base-neck rim. The neck height is almost equal to the height of the base, which is well rounded and penetrated by bundles of Sharpey’s fibres aligned in concentric circles over the base surface.

Cheiracanthus murchisoni scale morphology. (1)-(11) NMS G.2019.3.6 from Gamrie: (1)-(6) scanning electron microscope images of scale patch NMS G.2019.3.6.35 removed from midflank of counterpart; (7)-(11) drawings of scale crowns from: (7) behind the head; (8) midbody; (9) near the dorsal fin spine; (10) in front of the tail; (11) on the tail. (12)-(15) Scanning electron microscope images of detached midbody? scales of NMS G.2019.9.29 from Cromarty Sutors: (12) NMS G.2019.9.29.11.5, crown view; (13) NMS G.2019.9.29.11.2, laterocrown view; (14) NMS G.2019.9.29.11.19, lateral view; (15) NMS G.2019.9.29.11.1, anterolateral view. (16), (17) NMS G.2019.9.29.12 from Cromarty Sutors: (16) NMS G.2019.9.29.12.3, base showing concentric lines of Sharpey’s fibre bundles; (17) NMS G.2019.9.29.12.2, patch of articulated scales, crown view. (18), (19) ventral mid-body squamation patches of NMS G.2019.9.31.2 from Den of Findon. (20) NMS G.2019.9.30.2 from Edderton, scale in squamation patch. Scale bars equal 1.0 mm in 1-3, 0.3 mm in (4)-(11); (7)-(11) all at same scale, (17), (18), (20); 0.1 mm in (12)-(16), (20). Burrow et al. (2020).
Burrow et al.'s observations of the histological structure of the scales of Cheiracanthus murchisoni indicate that all growth zones, except the embryonic one, show ridges. As in all acanthodiforms, crown growth zones are superposed. Each zone is slightly wider in the neck than in the upper crown plane, and all zones outside the embryonic one are of similar width. Canals extend upwards and down into the base from the lacunae in the embryonic zone. Eight radial canals extend from openings close to the neck-base rim towards the embryonic zone, with ascending canals rising up in the middle of each growth zone. The four posterior canals open out through small calibre canals on each side of the two neck protuberances. Ascending canals branch and form an irregular anastomizing network. Near the anterior edge of the scales, canals from the network in the neck turn sharply back horizontally, and are interconnected by a horizontal ring canal completely encircling each growth zone in the inner zones, and only along the anterior edge in the outer growth zones. Lacunae are formed at the junction of the canals. The horizontal canals lie under the grooves on the crown surface. Along the posterior sides of scales, the anastomising networks forming the ascending canals gradually turn posteriorly and peter out into delicate dentine tubules reaching towards the surface of each growth zone. The central depression in the posterior surface of the crown of some scales appears to be related to a lower density of the networks of canals in this area of the scale. The central area of the upper plane in each crown growth zone is enameloid, with no dentine tubules visible. The base is formed of acellular bone lamellae. Sharpey’s fibre bundles and canals of Williamson extend through the base. Scales in the proximal area of fin webs have a similar profile to the body scales, decreasing in size distally to scales with a low flat base with a thin elongate crown. Thin fin rays underly the fin scales of each side of the web, apparently correlated one to one with the scales. Being clearly overlain by normal scales, they are surely directly comparable with the endoskeletal ceratotrichia of Chondrichthyans rather than the dermal lepidotrichia of Actinopterygians.

Cheiracanthus murchisoni scale histology. (1)-(5), (9)-(11): thin sections of scales from NMS G.2019.3.6 from Gamrie: (1) NMS G.2019.3.6.20, slightly oblique vertical section of mid-body scale; (2) NMS G.2019.3.6.32, vertical transverse section of ventral scale in the shoulder girdle region; (3) NMS G.2019.3.6.19, vertical transverse section of scale behind the head; (4) NMS G.2019.3.6.33, vertical longitudinal section of two contiguous scales in the midbody flank region; (5) NMS G.2019.3.6.36, subhorizontal section through base and lower crown; (9)-(11) NMS G.2019.3.6.28, vertical longitudinal section through fin web and body scales near pectoral fin spine: (9) distal fin scales; (10) fin web and body scales; (11) fin web scale and fin rays (ceratotrichia). (6) NMS G.2019.9.29.5 from Cromarty, vertical oblique section. (7) NMS G.2019.9.29.10 from Cromarty Sutors, vertical transverse section through posterior crown; (8) NMS G.2019.9.33.6, from Jessie Port, vertical transverse section. Abbreviations: ac, ascending canals; bl, base lamellae; cir, circular canals; cW, canals of Williamson; en, enameloid; ez, embryonic zone; fr, fin ray; lac, canal lacunae; p, protuberance; rc, radial canal; Sf, Sharpey’s fibre bundles. Arrows indicate anterior. Scale bars equal 0.1 mm. Burrow et al. (2020).
Cheiracanthus grandispinus is a species of Cheiracanthus with the Meckel’s cartilage very deep at the posterior, the maximum depth being about one quarter of the length; the scapula narrowest midshaft and widening dorsally and ventrally; the branchiostegal rays broad, long, and ornamented with longitudinal ridges, with thinner rays dorsally, bending down at their posterior end; endoskeletal gill bars with short lateral projections; scale crown with marked smooth rim anteriorly, fan-shaped arrangement of ridges extending the length of crown, with narrow ridges curving away from a deep central groove posteriorly.
Presumed adult specimens of Cheiracanthus grandispinus are over 200 mm long, with the largest estimated to be up to 350 mm long.
The jaw cartilages are similar to those of Cheiracanthus murchisoni, differing mainly in the greater relative depth of the Meckel’s cartilage posteriorly. The maximum depth of the palatoquadrate is abouy twice the maximum depth of the Meckel’s cartilage. The fossa towards the posterior end for insertion of the adductor muscle extends about half the length of the jaw. The preglenoid process is quite rounded, and the articular cotylus is a narrow embayment of the usual form in Acanthodians. The lateral face of the palatoquadrate has an extrapalatoquadrate ridge identical to that in Cheiracanthus murchisoni, and also has a vacuity in the same position as in Cheiracanthus murchisoni. The branchiostegal rays, with about eight below the jaw articulation and about 15 above, are quite broad and long, extending from the ventral edge of Meckel’s cartilage to about midway up the posterior edge of the palatoquadrate. The rays are ornamented with parallel longitudinal ridges. Long thin rays above the main branchiostegals are oriented obliquely, angled back and down with a downward turn at their posterior end. These are much more fragile than the branchiostegal rays and are rarely preserved. Numerous short, thin gular rays are present below the jaw line.

Cheiracanthus grandispinus head and branchial region, morphology. (1)-(2) NMS G.1966.40.24 from Lethen Bar. (3) NMS G. FR1603 from Achanarras Quarry, Caithness. (4) NMS G.2002.26.1481 from Achanarras Quarry. (5) NMS G.1903.130.19 from Achanarras Quarry, dorsoventrally flattened. (6) NHMUK PVP.1363 from Orkney. Abbreviations: addf, adductor muscle fossa; artc, articular cotylus; br, branchiostegal rays; cr.epq, extrapalatoquadrate ridge; gr, gular rays; Mc, Meckel’s cartilage; pdbr, posterodorsal branchiostegal rays; pq, palatoquadrate; pqv, palatoquadrate vacuity; pregl, preglenoid process. Anterior to right in (1)-(3), (6), to left in (4)-(5). Scale bars equal 10 mm. Burrow et al. (2020).
Notable histological features revealed by the thin sections through the branchial region are stacks of short mineralised endoskeletal elements that we identify as gill bars, or branchial rays, which we surmise to have extended out from the (unmineralised) branchial arches. Many of the gill bars in Cheiracanthus grandispinus have projections presumed to act as gill rakers on both sides. This arrangement, with branchial rays extending out from the arches, is like that in modern Chondrichthyans, rather than in the well-studied Acanthodiform Acanthodes, which has separate robust gill rakers attached directly on the branchial arches, and apparently lacks branchial rays. No mineralised gill bars were seen in the sacrificed specimen of Cheiracanthus murchisoni. The jaws are composed of blocky and globular calcified cartilage, as in Cheiracanthus murchisoni.

Cheiracanthus grandispinus head and branchial region, histology. (1)-(3) NMS G.2019.9.8: (1), (2) NMS G.2019.9.8.8 series of endoskeletal branchial rays, some showing gill raker projections; (3) NMS G.2019.9.8.14 calcified cartilage blocks; (4) NMS G.2019.9.35.1, calcified cartilage of Meckel’s cartilage? Abbreviations: gb, gill bars; gbp, gill bar projections; gcc, globular calcified cartilage; s, scale. Scale bars equal 0.1 mm. Burrow et al. (2020).
The scapulocoracoid is more robust than that of Cheiracanthus murchisoni, with smallest diameter near the middle of the scapular shaft, and widening towards the dorsal end The coracoid height is equal to or greater than scapular height. A separate procoracoid was not preserved in any of the specimens examined. The spines show a similar morphology to those of Cheiracanthus murchisoni, but the pectoral fin spines are slightly more robust. The dorsal spine is straight, positioned slightly behind the level of the pelvic spines, and has a basal cartilage.

Cheiracanthus grandispinus pectoral region, spines morphology. (1) USCP F00130a from Achanarras, whole fish. (2) NMS G.FR1810 from Achanarras, scapulocoracoids and pectoral fin spines. (3) NMS G.2019.9.12, isolated scapulocoracoid from Achanarras. (4) Disarticulated incomplete Fish NMS G.2019.9.10.1 from Cromarty, ?two pectoral, one median (?dorsal), one pelvic spine. Abbreviations cor, coracoid; fs, indeterminate fin spine, probably a pelvic; median fs, dorsal or anal fin spine; pect.fs, pectoral fin spine; pect. fw, pectoral fin web; sc.s, scapular shaft. Anterior to right. Scale bars equal 50 mm in (1), 10 mm in (3), (4), 2 mm in (2). Burrow et al. (2020).
Relative heights and widths of the leading edge ridge and body of spine vary depending on the spine position. In paired spines, the leading edge ridge is higher than wide; pectoral spines have an elongate leading edge ridge, and pelvic spines have a wide leading edge ridge. In median fin spines, this ridge is wider than high. All spines, like those of other Cheiracanthus species, have a thin enameloid outer layer preserved on the leading edge ridge and the ‘shoulders’. Spines have a wide central cavity and lack an accessory pulp canal. Osteodentine forms most of the spine, with dentine tubules mostly only visible branching out towards the surface of the spine from the outermost longitudinal canals. Longitudinal and interconnecting canals extend the length and breadth of the spine, with smaller canals leading off to open out in the sulcus between the leading edge ridge and the ‘shoulders’ of the spines, as well as into the central cavity. Canals radiate out from the longitudinal canals in the leading edge ridge. Denteons are formed around the canals in the exserted part, but not in the inserted part. The inner lamellar layer is present proximal to the closure of the central cavity, and increases in thickness distally. Branching dentine tubules extend through each of the lamellar layers, perpendicular to the inner surface. In the Achanarras preservation type, a clear boundary is visible between the denser tissue of the posterior face of the spine and the osteodentine and inner lamellar layers of the main body of the spine. Remnants of a separate thin layer of tissue lining part of the central cavity are preserved towards the proximal end in some of the spines.
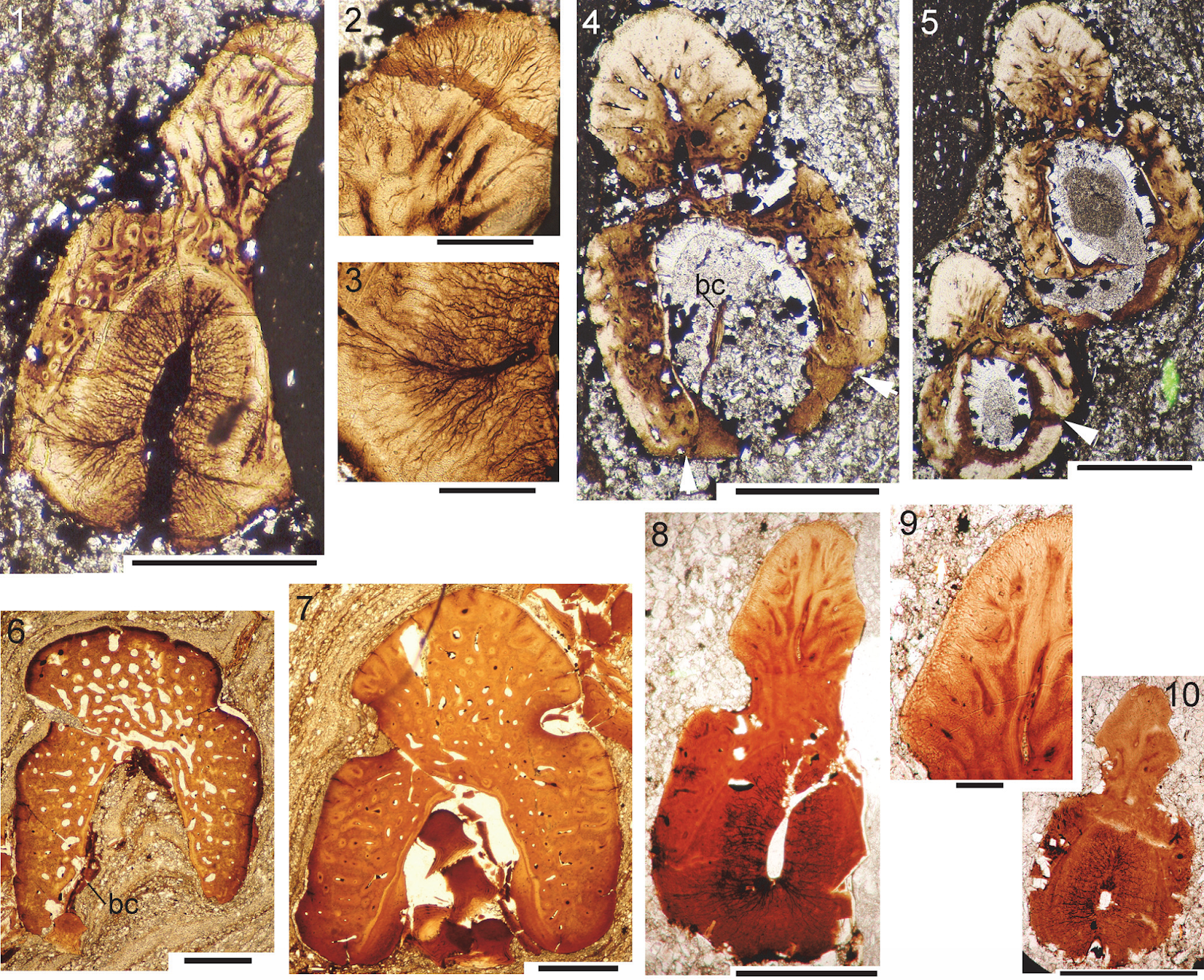
Cheiracanthus grandispinus spines histology. (1)-(5) NMS G.2019.9.7 from Achanarras: (1)-(3) NMS G.2019.9.7.21, thin section of distal end of pectoral spine, with closeups of the structure of the leading edge ridge and boundary between the main spine osteodentine and the areas bounding the trailing edge groove; (4) NMS G.2019.9.7.11, TS pelvic spine; (5) NMS G.2019.9.7.12, TS through both pelvic fin spines. (6), (7) NMS G.2019.9.8 from Jessie Port, median spine, ?anal: (6) TS NMS G.2019.9.8.8 at proximal end of exserted part; (7) NMS G.2019.9.8.15, towards mid spine. (8)-(10) NMS G.2019.9.35.1-7 from Achanarras, pectoral spine: (8), (9) NMS G.2019.9.35.2, midspine; (10) NMS G.2019.9.35.3, more distal. Abbreviation: bc, basal cartilage. Scale bars equal 0.5 mm in (1), (4)-(8), (10), 0.1 mm in (2), (3), (9). Burrow et al. (2020).
Scales of larger Fish (200 mm or more long) are easily distinguishable from scales of other species of Cheiracanthus, with a fan-shaped arrangement of crown ridges anteriorly, and with the ridges curving away from a deep central groove posteriorly in the midline. Even in the fin web scales, the same fan-shaped pattern is visible. The crown is up to 1.5 mm long on flank scales and usually longer than wide. The crown ridges are narrow but vary slightly in width, apparently randomly, and extend the whole length of the crown. Particularly on worn scale crowns, the ornament more resembles grooves cut into the crown, rather than ridges developed above the crown plane. Broad low longitudinal swellings line the median groove. Extra ridges are developed laterally in the posterior half of the crown, usually at a slightly lower level than the central crown. A broad smooth rim is present in front of the ridges along the anterior edge. In scales from smaller, presumed juvenile Fish, and in the older growth zones, crown ridges are more radially arranged, extra lateral ridges are lacking, and the postero-lateral edges are sometimes serrated. This morphology is visible on primordial scale crowns exposed through flaking off of overlying growth zones. The surface of the crown in all scales is convex both longitudinally and side to side. The neck and base are developed similarly to those in Cheiracanthus murchisoni, with a neck that is as deep or deeper than the base, and a pair of protuberances posteriorly.

Cheiracanthus grandispinus scale morphology. (1)-(3), (5)-(9), light microscope images; (4), (10)-(18) scanning electron microscope images; midbody scales unless detailed otherwise. (1) NMS G.1891.92.317 from Lethen Bar. (2) NMS G.2019.9.8 from Jessie Port, Ross and Cromarty. (3), (4) USCP F00115b from Edderton. (5) NMS G.2019.9.9 from Achanarras, primordial scale revealed after abrasion/flaking of overlying crown growth zones. (6)-(9) NMS G.2019.9.10.1 from Cromarty: (6) mid-body scales; (7)-(9) pectoral fin web: spine and web; (8) distal scales; (9) scales near fin base. (10)-(12) NMS G.2019.9.11 from Marwick: (10) view of crown before removal of scale from matrix; (11) anterocrown view; (12) posterior view. (13) NMS G.2019.9.3.27.1 from Gamrie Den of Findon, laterocrown view. (14)-(18) NMS G.2019.9.13.8 from Cromarty, Eathie to Navity: (14) NMS G.2019.9.13.8.16, anterocrown view; (15) NMS G.2019.9.13.8.2 pair of scales in posterocrown view; (16) NMS G.2019.9.13.8.8, laterocrown view; (17) NMS G.2019.9.13.8.6, lateral view; (18) NMS G.2019.9.13.8.17, posterobasal view. Abbreviations: p, neck protuberance; ps, primordial scale. Arrows indicate anterior. Scale bars equal 0.5 mm in (1)-(9), 0.2 mm in (10)-(18). Burrow et al. (2020).
In many aspects, the histological structure of the scales is similar to that of Cheiracanthus murchisoni. The embryonic zone (primordial scale) has large lacunae interconnected by short narrow canals, with other short canals extending upwards and down towards the base cone. The crown surface of the primordial scale is concave in longitudinal and transverse sections. In younger zones the surfaces become more and more convex both longitudinally and transversely. The deep median longitudinal groove extends through all the zones in the posterior half of the scale.

Cheiracanthus grandispinus scale histology. (1) NMS G.2019.9.3.9 from Gamrie, vertical longitudinal section through posterior protuberance. (2), (9), (12) NMS G.2019.9.8 from Jessie Port, Ross and Cromarty: (2) NMS G.2019.9.8.14, anterior crown vertical transverse section; (9) NMS G.2019.9.8.10, crown vertical transverse section; (12) NMS G.2019.9.8.13, crown horizontal section. (3) NMS G.2019.9.35.6, from Achanarras, midscale vertical transverse section. (4), (5), (10), (11) from Achanarras: (4) NMS G.2019.9.2.4, posterior half of scale, vertical transverse section; (5) NMS G.2019.9.2.2, vertical longitudinal section; (10) NMS G.1893.107.9.2, horizontal section through crown base; (11) NMS G.1893.107.9.2, horizontal section through midcrown. (6)-(8) NMS G.2019.9.10 from Cromarty: (6) NMS G.2019.9.10.3, off-centre vertical longitudinal section; (7) NMS G.2019.9.10.5, vertical transverse section; (8) NMS G.2019.9.10.9, oblique vertical section through side of scale. Arrows indicate anterior. Scale bars equal 0.1 mm. Burrow et al. (2020).
As in Cheiracanthus murchisoni, eight regularly spaced, radial canals extend through the lower crown, with anastomizing networks of ascending canals running up in each crown growth zone. The junction between the posterior radial and ascending canals shows a wide lacuna in the younger growth zones. Anteriorly, the ascending canals are interconnected in a ring canal consisting of short, arched tubules before turning back horizontally. In young individuals and juvenile growth stages, each groove between the more lateral ridges is usually underlain by only one horizontal canal, and the ridges appear gently rounded in cross section. With growth of the scales the younger ridges show a more flat-topped shape, and the grooves are underlain by two to four canals which sometimes intertwine. More posteriorly, regularly placed dentinal tubules run off the canals into the ridges, sometimes forming a network between adjacent ridges. Enameloid fills most of the upper plane in each crown growth zone, with longitudinal dentine tubules near the base of each zone. Sharpey’s fibre bundles in the base are wider than those of Cheiracanthus murchisoni, so the isopedin-like layering of the sheets of fibres appears coarser than in that species. Otherwise, the bases resemble those of Cheiracanthus murchisoni.

Cheiracanthus grandispinus comparison of scale structure in
different sized fish. (1) NMS G.2019.9.5.2, from Achanarras, small Fish,
about 10 cm long, vertical transverse section through scale anterior.
(2) NMS G.2019.9.6.1, from Achanarras, medium sized fish less than 30 cm
long, vertical transverse section through scale anterior. (3) NMS
G.2019.9.8.12, from Jessie Port, medium sized Fish, vertical transverse
section through scale anterior. (4) NMS G.2019.9.9.14, from Achanarras,
Fish 30 cm or longer, vertical transverse section through scale
anterior. (5) NMS G.2019.9.9.7, from Achanarras, Fish 30 cm or longer,
vertical longitudinal section. NMS G.2019.9.35.4, medium sized
incomplete Fish, vertical section of scale showing Sharpey’s fibre
bundles through base. Scale bars equal 0.1 mm. Burrow et al. (2020).
The number of crown growth zones increases as the size of the Fish increases. Up to seven crown growth zones were noted in scales of the smallest Fish, which was about 100 mm long, and up to 15 in the largest fish sampled, which were over 300 mm long.

Cheiracanthus grandispinus head and branchial region reconstruction. Burrows et al. (2020).
Cheiracanthus latus is a species of Cheiracanthus with a large deep tail, at its greatest depth about one-third the length of the Fish; deep palatoquadrate with maximum depth about 2.5 times maximum depth of the Meckel’s cartilage; Lunate dorsal circumorbital bone; dorsal branchiostegal rays very short and slender with a distinct bend downwards on their anterior ends; long, thin rays above the main branchial cover; scale crowns with two broad median ridges edging median longitudinal sulcus with an oval pit in the posterior half; posterior crown with serrated margin.
The most obvious feature distinguishing Cheiracanthus latus from Cheiracanthus murchisoni and Cheiracanthus grandispinus is the large caudal fin. The dorsal spine is slender and straight, positioned midway between the levels of the pelvic and anal spines. The anal spine is also slender and straight, and about 80% the length of the dorsal spine. The pelvic spines show a very slight curvature, and are about 65% the length of the dorsal spine. The pectoral spines are a similar length to the dorsal spine and slightly curved.
The head of Cheiracanthus latus is very similar to that of Cheiracanthus murchisoni. The only difference previously noted is that the dorsalmost branchiostegal rays in Cheiracanthus latus are shorter, slenderer, and the anterior ends turn sharply ventrally. Burrow et al. note that the main branchiostegal rays are ornamented with thin parallel longitudinal ridges, however, this ornament is rarely seen as the rays usually fracture centrally when nodules are split. NHM P43273 is a rare specimen where it is visible. They also note that Cheiracanthus latus has distinct circumorbital bones with the most dorsal one extending about one-fifth the circumference of the orbit.

Cheiracanthus latus head and branchial region, morphology. (1)-(2) NHM PV P 3253 from Tynet Burn. (3) NMS G.1891.92.320 from Gamrie. (4) NHMUK PVP43273 from Tynet Burn, branchial region with ridges visible on lowest branchiostegal plates. Abbreviations: br.l, left branchiostegal plates; br.r, right branchiostegal plates; cop, circumorbital plate; gr, gular rays; Mc.l, left Meckel’s cartilage; Mc.r, right Meckel’s cartilage; pect.fs, pectoral fin spines; pq.l, mineralised cartilage along ventral edge of left palatoquadrate; proc, procoracoid; sc, scapulocoracoids; tess, head tesserae. Scale bars equal 10 mm. Burrow et al. (2020).
Thin sections show that the jaws and most other endoskeletal structures in these regions are mineralised as calcified cartilage with the same basic varieties as in Cheiracanthus murchisoni. The jaws are mostly a single layer of contiguous calcified cartilage blocks, and the branchial arches comprise a more irregular consolidated globular calcified cartilage forming cylinders filled with calcite that has presumably replaced the uncalcified cartilage core. The endoskeletal branchial arches show a similar structure as in Cheiracanthus murchisoni, but the walls are thinner and appear to comprise only one layer. As the Cheiracanthus latus Fish sacrificed for thin sectioning was larger than the Cheiracanthus murchisoni one, it is unlikely that the difference in thickness is related to the size of the Fish. The dermal branchiostegal plates and smaller gular rays are formed of a dense lamellar bone; the ornament layer on the branchiostegal rays comprises sharp crested ridges. Small mono- and multicuspid denticles are preserved around the branchiostegal rays. Blocks of short, thin mineralised elements within the branchial regions are interpreted as the endoskeletal gill bars some show short projections. Flat or concave based dermal tesserae from the head region have a lamellar base and areal-growth odontodes formed of acellular layered dentine.

Cheiracanthus latus head and branchial region, histology, NMS G.2019.3.3. (1), (2) thin section NMS G.2019.3.3.3: (1) ?Meckel’s cartilage; (2) closeup of calcified cartilage blocks; (3), (4) NMS G.2019.3.3.5, indet. jaw cartilage and branchial arches; (5) NMS G.2019.3.3.2, branchiostegal plate in oblique section, body scales, branchial denticles, indet. cartilage; (6) NMS G.2019.3.3.4, body scales, gill bars; (7), (8) NMS G.2019.3.3.17: (7) jaw cartilage, head tesserae, gill bars, branchiostegal plates; (8) head tessera vertical section, stellate mineralisation artifacts. Abbreviations: ba, branchial arch; br, branchiostegal plate; ccb, calcified cartilage blocks; d, denticle; gb, gill bar; gcc, globular calcified cartilage; gz, growth zones in crown; s, body scale; t, tessera. Scale bars equal 1.0 mm in (1), (3), (5), (7), 0.1 mm in (2), (4), (6), (8). Burrow et al. (2020).
The scapulocoracoid of Cheiracanthus latus closely resembles that of Cheiracanthus murchisoni, with a slender tapering scapular shaft. Burrow et al. note that there is a marked delineation between the scapula and the coracoid ossifications.
None of the fin spines are deeply inserted, the dorsal fin spine has a basal bone; this is a short D-shaped mineralised cartilage. The pulp cavity is open for about half the length of the spine. The pectoral spines have a smooth leading edge ridge, deep lateral groove, and thin longitudinal ridges along the side, usually best developed more proximally. A single row of pores opens out in the groove. They have a short insertion, and a strip lacking dentine extending along the upper side of the spine, where the scapulocoracoid abutted. Like the dorsal spine, the anal and pelvic spines have a smooth leading edge ridge separated from the smooth sides by a deep groove.

Cheiracanthus latus pectoral region and spines, morphology. (1)-(3) QMF60004 from Tynet Burn: (1) left and right scapulocoracoids, white arrows indicate separation between scapula and coracoid; (2) close up of right coracoid shape (dotted line); (3) dorsal fin spine insertion and basal cartilage, white arrow indicates insertion/exertion boundary. (4)-(6) NMS G.2018.28.26 from Tarrel Bay: (4) left and right pectoral spines as collected; (5) closeup of thin ridges on lateral or medial? side of right spine; (6) insertion area of right spine. (7) NMS G.1968.19.18, anal and pelvic spines. Abbreviations: a.fs, anal fin spine; bc, basal cartilage; calc, calcite infill; pelv.fs, pelvic fin spines. Scale bars equal 5.0 mm in (1)-(3), (6), 20 mm in (4), (7), 0.5 mm in (5). Burrow et al. (2020).
The scapulocoracoid is formed of a dense lamellar bone around a central cavity. The histological structure of the fin spines is very similar to that of Cheiracanthus murchisoni and Cheiracanthus grandispinus, the main difference being that the lateral groove is not as deeply incised. The pectoral spines have a very thin enameloid layer on the leading edge ridge and the ‘shoulders’, with most of the spine formed of osteodentine. Spines lack an accessory pulp canal. Pores leading from the vascular canals open out into the lateral groove. Dentine tubules of the osteodentine layer are usually only visible near the surface of the anterior ridge, oriented perpendicular to the surface and running in to the more or less radially arranged vascular canals. The inner lamellar layer is very thin proximally, thickening distally, and is penetrated by fine branching dentine tubules. The vascular canals and the central pulp cavity become almost closed by lamellar infilling towards the tip of the spine, but canals still extend from the cavity through the dentine. The inserted part of the spine lacks an outer enameloid cover and dentine tubules, and is composed of a vacuous osteodentine or bone. The thin longitudinal ridges on the sides of the spines are low, sharp-crested, and best visible in the proximal part of the spine.

Cheiracanthus latus pectoral region and spines, histology. (1)-(6) NMS G.2019.3.31: (1) slice II before grinding (corresponds to sections (6), (7)), arrow indicates dorsal direction; (2) NMS G.2019.3.3.7, scapulocoracoid, long section mid-shaft; (3) NMS G.2019.3.3.12, thin section of pectoral spine at insertion/exertion boundary; (4) NMS G.2019.3.3.10, thin section of spine close to insertion; (5) NMS G.2019.3.3.6, close up of leading edge ridge structure of pectoral spine, midspine; (6) NMS G.2019.3.3.2, thin section of pectoral spine near the tip. (7)-(10), NMS G.2018.28.26, thin sections of pectoral spines: (7) 2018.28.26.9, close to insertion; (8) 2018.28.26.7, midspine; (9), (10) 2018.28.26.5, towards tip, with closeup of infilling dentine. (11)-(14) NMS G.2019.3.7 thin sections of pectoral spine, near insertion, and flange of scapulocoracoid: (11) NMS G.2019.3.7.8; (12) NMS G.2019.3.7.9; (13)-(14) NMS G.2019.3.7.10, outer side of spine with thin sharp ridges. Abbreviations: c.cav, central cavity; ccb, calcified cartilage blocks; dt, dentine tubules; en, enameloid; l.pect.fs, left pectoral fin spine; min, stellate mineralisation; ost, osteodenteon; pect.fs, pectoral fin spine; r, ridges; s, scale; sc, scapulocoracoid. Scale bars equal 10 mm in (1); 0.1 mm in (2), (5), (10); 0.5 mm in (3), (4), (6), (7)-(9), (11)-(14). Burrow et al. (2020).
The mid-flank scales of Cheiracanthus latus range in size from 0.25 mm wide on small Fish, about 130 mm long, about 0.5 mm wide on medium sized Fish, to 1.0 mm wide on large fish. The scale crown has a marked smooth rim along the anterior edge. The ornament over the rest of the crown comprises two prominent, broad ridges extending back on either side of a median sulcus, and narrower, sharp-crested ridges on both sides of the median ridges extending back towards the serrated posterolateral edges of the crown. A few short ridges are developed in the anterior part of the median sulcus, and an oval pit is developed in the posterior half of the scale within the sulcus. The neck and base of the scales closely resemble those of Cheiracanthus murchisoni in general profile and relative heights.

Cheiracanthus latus squamation, light microscope images. (1), (2) QMF60004 from Tynet Burn: (1) midflank scales; (2) caudal peduncle, scale impressions. (3), (4) NMS G.2018.28.26 from Tarrel Bay. (5)-(7) NMS G.2019.3.7 from Den of Findon, Banffshire (8) NMS G.2019.3.3 from Jessie Port. (9) NMS G.1870.14.145 from Cromarty. (10) NMS G.2019.9.17 from Geanies Point. Abbreviation: p, pit. Arrows indicate anterior. Scale bars equal 0.5 mm.
In most histological features the scales of Cheiracanthus latus resemble those of Cheiracanthus murchisoni, other than the development of a wide midline channel extending the length of the crown, which usually deepens to form an oval pit just posterior to the centre of the scale. The ridges extending back from the anterior edge are sharp-crested near this edge, becoming rounded more posteriorly, and often fading out past the posterior median pit. The crown is formed of up to 10 growth zones, which show that the wide central pit/channel was present at all stages from the embryonic/primordial scale. As in Cheiracanthus murchisoni, the primordial scale (roughly 60 μm long and 70 μm wide) contains wide lacunae connected by short canals with offshoots extending down into the top of the basal cone. Each growth zone has its own system of anastomizing ascending canals, building an intricate network especially in the posterior side of the scales. The anterior areas of the crown growth zones are developed similarly to those of Cheiracanthus murchisoni, but differ in that the horizontal canals in the upper crown leading back from the semicircular ring canal are arranged much less regularly, and the lateral canals are better developed than the central canals. This central area is delimited by two strongly developed horizontal canals, which extend further posteriorly than the other canals, in the larger ribs that flank the central pit. The horizontal canals are always situated in the grooves between crown ridges. In the posterior half of the crown, the ascending networks of canals gradually turn from a vertical into a more horizontal direction. The enameloid layers in the crown growth zones fill most of the depth of each zone in the ridges flanking the central pit, diminishing in depth laterally and disappear about halfway between the midline and the lateral edge. In the midline of the crown, enameloid layers are only developed anterior to the central channel and are completely lacking along most of the length. Fin web scales distal to the fin base differ markedly to the flank scales in their general shape, having a flat or concave base, deep neck, and crown plane that is concave both anteroposteriorly and side to side.

Cheiracanthus latus squamation, scanning electron miccroscope images. (1)-(5) NMS G. 2019.3.7.21 from Den of Findon: (1) NMS G. 2019.3.7.21.2, crown view; (2) NMS G. 2019.3.7.21.5, crown view; (3) NMS G. 2019.3.7.21.13, anterior view; (4) NMS G. 2019.3.7.21.7, laterocrown view; (5) NMS G. 2019.3.7.21.14, laterocrown view. (6)-(7): NMS G.2019.14.9.2 from Geanies Point: (6) NMS G.2019.14.9.2.8, anterocrown view; (7) NMS G.2019.14.9.2.5, crown view. (8)-(10): NMS G.2019.14.4.2 from Hilton or Cadboll: (8) NMS G.2019.14.4.2.15, crown view; (9) NMS G.2019.14.4.2.5, crown view; (10) NMS G.2019.14.4.2.16, laterocrown view. Abbreviations: p, pit; pr, neck protuberance. Scale bars equal 0.2 mm. Burrow et al. (2020).
Although some aspects of the morphology of Cheiracanthus murchisoni and Cheiracanthus latus have been described previously, Burrow et al. provide the first detailed description of Cheiracanthus grandispinus, showing that it is a valid species, characterised by the robustness of its fin spines and scapulocoracoid, and the distinctive fan-shaped ornament ridges on the scale crowns.

Cheiracanthus latus scale histology. (1) NMS G.2019.9.21.5, vertical transverse section, anterior half of scale. (2) NMS G.2019.9.19.2, vertical transverse section, anterior half of scale. (3) NMS G.2019.9.20.3, vertical transverse section, midscale. (4), (5) NMS G.2019.3.3.14, vertical transverse section, midscale, with closeup of central region. (6) NMS G.2019.9.21.3, vertical transverse sections of two scales, posterior half of scale. (7) NMS G.2019.3.3.12, vertical transverse section towards posterior end of scale. (8) NMS G.2019.3.7.6, subhorizontal section through anterior base and posterior crown. (9) NMS G.2019.3.7.6, vertical oblique section through posterior crown and base. (10) NMS G.2019.9.19.7, horizontal section low in crown. (11) NMS G.2019.9.22.5, horizontal section high in crown. (12), (13) NMS G.2019.9.23.7, section through fin web scales including tiny distal scales with a deep neck and flat base, closeup of distal fin web scales in (13). Arrows indicate anterior. Scale bars equal 0.1 mm in (1)-(11), (13), 0.5 mm in (12). Burrow et al. (2020).
One of the features, which we describe for the first time in these three Cheiracanthus species, is the presence of denticles and gill bars in the branchial region, and tooth-like elements in the orobranchial region. The latter are smaller than those noted in the Cheiracanthid Homalacanthus concinnus, and only rarely visible on articulated Cheiracanthus specimens, but are clearly seen in the serial sections, which show that they are formed of dentine like the ones in Homalacanthus concinnus. It seems unlikely that they are typical gill rakers, because they are only visible in the mouth region of articulated and sectioned Cheiracanthus specimens. It is perhaps surprising that the elements have been ignored since the original description of Cheiracanthus latus, but they are difficult to see in most specimens.

Cheiracanthus latus head and branchial region reconstruction. Burrow et al. (2020).
Burrow et al. have also identified the bi-partite structure of the endoskeletal pectoral girdle, with a clear demarcation visible between the scapular and coracoid parts. Another notable observation relates to the D-shaped cartilages often mineralised at the base of the median fin spines: thin sections show that this cartilage extends into the pulp cavity of the fin spine as a thin lining, reminiscent of the basal cartilages in both extant and extinct Chondrichthyans.
As well as the differences between the scales of the Cheiracanthus species, Burrow et al. have identified other morphological characters to distinguish between them: the scapular shaft is relatively more robust in Cheiracanthus grandispinus and expands dorsally, whereas it narrows in Cheiracanthus murchisoni and Cheiracanthus latus; the pectoral fin spine in C. murchisoni has thin longitudinal ridges ornamenting the sides, which are smooth in Cheiracanthus grandispinus and Cheiracanthus latus; the height of the leading edge ridge on fin spines of Cheiracanthus grandispinus is more than a third the total height of the spine, whereas it is a quarter or less the total height in Cheiracanthus murchisoni and Cheiracanthus latus; Cheiracanthus latus has a semi-lunar circumorbital plate above the orbit.

Reconstruction of a whole Cheiracanthus latus in lateral view. Burrow et al. (2020).
Burrow et al.'s investigations have revealed new details of the structure of the endoskeletal tissues in Cheiracanthus, in particular the composition of the jaws and branchial arches. The endocranium was previously considered to be preserved as perichondral bone, but serial thin sections of all three species Cheiracanthus murchisoni, Cheiracanthus grandispinus, and Cheiracanthus latus (as well as the recently described new species Cheiracanthus peachi) have shown that when the neurocranium and/or splanchocranium is mineralised, it is mostly as a single layer of calcified cartilage, often formed of contiguous blocks which are sub-rectangular in vertical section. In addition, some of the branchial bars are mineralised as a two-layered, consolidated calcified cartilage. Burrow et al. note that these two forms of calcified cartilage also occur in the same elements in the Diplacanthiform Diplacanthus crassisimus. This co-occurrence could have interesting connotations from a phylogenetic perspective. Cheiracanthids are united with other Acanthodiforms based primarily on Acanthodids, Mesacanthids, and Cheiracanthids only having one dorsal fin spine. However, cheiracanthids differ from the other two families in having ornamented scale crowns, jaw cartilages mineralised as single units, lack of mandibular bones, and as we have shown here, an endoskeleton (other than the pectoral girdle) mineralised as calcified cartilage, not perichondral bone, and oral tooth-like elements. As the endoskeletal structure becomes better known and characterised in Acanthodians and other stem Chondrichthyans, their interrelationships will perhaps become clearer than cladistic analyses have so far deduced, with proposed clades within the group generally poorly supported in those analyses.
Burrow et al. also note that some of the Acanthodian species found in the Scottish Middle Devonian appear earlier in deposits further east. The taxa include Cheiracanthus latus and Diplacanthus crassisimus, which are both found in upper Emsian deposits of the Baltic region and Belarus. This stratigraphic distribution indicates that these species evolved in the eastern regions and migrated west to the Orcadian Basin of Scotland in the Eifelian. It has been noted elsewhere that this westward migration of other classes of Fish occurred in waves during the Middle Devonian, with very little endemism in the Orcadian Basin.
See also...





Follow Sciency Thoughts on Facebook.