In 1916 the father and son dinosaur hunting team Charles and Levi Sternberg mounted an expedition to the Steveville Badlands of Alberta, Canada to collect dinosaur bones for the
Natural History Museum in London. The area had proven a bountiful source of dinosaur bones for previous expeditions, having first been worked by Lawrence Lamb in 1898, and has remained a centre for dinosaur hunting to this day, forming part of
Dinosaur Provincial Park, an area so rich in dinosaur bones that palaeontologists have attempted to erect stratigraphic biozones based upon them. Erecting biozones is a method of using fossils to date rocks; this uses fossils with known chronological ranges to establish the maximum and minimum age of rocks containing them; usually biozones are erected using shellfish, micro-organisms, pollen, rodents' teeth or other such small widespread fossils.
Alberta proved to be a productive area for the Sternbergs, but failed to impress the Museum; the then Keeper of Geology, Arthur Smith Woodward, described the fossils returned as 'rubbish'. Woodward was a powerful and influential figure at the time, though not a wise one. He gained a reputation for using his influence to marginalize promising younger palaeontologists who he saw as a threat to his status, and had an unscientific tendency towards self promotion, attaching himself to newsworthy discoveries and burying others. He was eventually discredited by the Piltdown forgery.
The items sent by the Sternbergs to Woodward did not contain large, complete fossil dinosaurs that would make good displays in the Museum, so they were stored away and forgotten, and never really assessed for their scientific interest. The Sternbergs were apparently unconcerned by this; they were veteran fossil hunters, and if the British Museum was not interested in their fossils, then there were plenty of other museums that were.
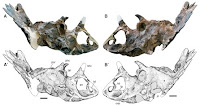
The new dinosaur has been given the name Sinops sternbergorum (Spinops meaning 'spineface' and sternbergorum in honor of its' discoverers. Sinops lived about 76 million years ago, in the Late Cretaceous. It is thought to have been about six meters long in life, and to have weighed about two tonnes. Like other Ceratopsid Dinosaurs it had a prominent facial shield, with several horns.
Artist's rendering of the skull bones of Sinops sterbergorum. Copyright Lukas Panzrin, courtesy of the Raymond M. Alf Museum of Paleontology.
This is where things got interesting. When Farke et al. examined the nasal horn of Sinops they realized that it erupted from a different part of the skull to that of better described Ceratopsid Dinosaurs. This was not entirely unexpected; it had been suggested of some other species, but in these cases the fossils had been imperfect and somewhat ambivalent. In the case of Sinops there was no doubt; the horn was sufficiently different from that of other ceratopsids that it must have arisen separately.
Sinops is a member of a group of ceratopsids called the Centrosaurinae. This group are considered basal or (misleadingly) primitive within the ceratopsid group. What this actually means is that the group divided from the other Ceratopsid Dinosaurs early in the family's history; it is not closely related to the most familiar ceratopsid, Triceratops.
The earliest known centrosaur derives from Late Cretaceous Oldman Formation of Alberta the is Albertaceratops nesmoi; it is about a million and a half years older than Sinops at roughly 77.5 million years old. It is known from a single skull and a few fragmentary bones from the rest of the body. Albertaceratops is bore long brow horns and a bony ridge on its nose where most ceratopsids have a horn, which suggests that it came from a lineage in which the horn had not yet arisen, this supports the idea that the nasal horn subsequently evolved in separate lineages. Albertaceratops also had two small, hook-like horns pointing outward at the top of it's crest.
An impression of Albertaceratops nesmoi by paleoartist Nobu Tamura.
The next centrosaur to appear in the fossil record is Centrosuarus, which first appears in the Oldman Formation of Alberta, about 65.5 million years ago, and continues into the overlying Dinosaur Park Formation. The genus after which the group is named. Centrosaurus had a large nasal horn, small brow horns and a row of small horns around the frill. Centrosaurus could be up to six meters long, but was not especially heavily built. There are two described species, C. apertus, in which the small brow horns curve upwards, and C. brinkmani, in which they curve to the side. It has a much better record than most other species, with several vast bone beads known. These are though to have formed where large numbers of individuals gathered around drying water holes and eventually perished.

The beds immediately overlying those containing Centrosaurus contain another species of centrosaur, Styracosaurus albertensis. This was a slightly shorter animal from tip to tail, but was much heavier built, reaching 1.8 m at the shoulder, and being loner in the body and shorter in the tail; it could probably weigh nearly 3 tonnes. Like Centrosaurus, Styracosaurus has been found in large bone beds, thought to have formed in mass death events around drying water holes in times of draught. Styracosaurus had a large, straight, nasal horn and almost non-existent brow horns. The edge of the frill bore a number of large horns rivaling the nasal horn in length. The nasal horn has not been found intact, so the exact size and development pattern of this horn is unclear.
A reconstruction of Styracosaurus albertensis by Nobu Tamura.
Diabloceratops eatoni lived 75 million years ago in what is now Utah. It is known only from a partial skull, with a pair of large spikes on the top of its neck frill, small brow horns and one or two small nasal horns (again this part of the skull is not well preserved). Diabloceratops is considered to be from a lineage that derived from the earliest centrosaurs separately; all other known centrosaurs are thought to be more clesely related to one-another than to Diabloceratops.
A reconstruction of Diabloceratops eatoni, by natural history illustrator Andrey Atuchin.
Einiosuarus procurvicornis is known from a number of bone beds from Montana, between 74.5 and 74 million years old. It reached about six meters in length, had a forward curving nasal horn, brow horns reduced to a pair of knobs, and a pair of horns on the top of its small neck frill.
From the same time in Montana came Rubeosaurus ovatus, known only from two partial skulls. Rubeosaurus appears to have had a large nasal horn, and at least two pairs of horns on the neck frill. Some juvenile fossils attributed to the genus Brachyceratops may also belong to this species.
Above these species in the same deposits in Montana, the species Achelousaurus horneri dates from about 74 million years ago, and is known from three skulls and a partial skeleton. It appears to have been about six meters in length, and to have had boney ridges in the place of the nasal and brow horns, though it did have a pair of horns on the top of the neck frill.
An illustration of Achelousaurus horneri by Sergey Krasovskiy.
The latest surviving genus of centrosaur appears to have been Pachyrhinosaurus. Two species of this are known from Alberta, P. lakustai dating from between 73.5 and 72.5 million year ago and P. canadensis from about 71.5 to 71 million years ago. A third species, P. perotorum has been found in Alaska, and is even younger, at between 70 and 69 million years old, making it the last known surviving centrosaur. Pachyrhinosaurus was about eight meters long when fully grown, and weighed about eight tonnes. The nose and brow horns were replaced by bony ridges, and the neck frill bore a single pair of small horns in P. canadensis and some specimens of P. lakustai. P. perotorum and some specimens of P. lakstai lacked these horns. In P. Canadensis and P. perotorum the brow ridges were close together, but in P. lakustai they were further apart.
Pachyrhinosaurus canadensis by Nobu Tamura.


 Regaliceratops peterhewsi: A new species of Chasmosaur from the Late Cretaceous of Alberta. Ceratopsids are among the most
distinctive and...
Regaliceratops peterhewsi: A new species of Chasmosaur from the Late Cretaceous of Alberta. Ceratopsids are among the most
distinctive and...

























