Bony-toothed Birds
(Odontopterygiformes: Pelagornithidae) are an extinct clade of large,
pelagic, volant Birds with a fossil record spanning
from the late Paleocene to the late Pliocene and a global distribution.
As their colloquial name suggests, the most obvious diagnostic
characteristic of this clade is the modification of the tomial crest of
the premaxillae, maxillae, and dentaries into a variety of tooth-like
bony projections that lack dental tissues (or homology to teeth). The
sizes and spacing of these projections vary across the clade but are
consistent within species, following a set sequence of large and small
pseudoteeth covered in life by the rhamphotheca. This pseudodentition,
along with hooked premaxillae and the presence of intraramal joints, has
been hypothesised to indicate dietary preferences for Fish or Squid
skimmed from the top of the water column. Pelagornithids and the extinct
Teratorns (Teratornithidae) from the Neogene and Quaternary are the
largest volant Birds known, and while the body sizes of Pelagornithids
vary, the majority of known specimens and species derive from
individuals considered large (3.5–4.5 m wingspan) and even giant (5–6 m
wingspan).
Though they have a nearly global distribution, Pelagornithid specimens from Antarctica are rare and limited to isolated elements; most of which derive from the Eocene sediments of the La Meseta and Submeseta Formations on Seymour (Marambio) Island, near the Antarctic Peninsula. The published specimens from these formations include two partial maxillae, one fragment of a humerus, one dentary fragment, and one distal tarsometatarsus.
In a
paper published in the journal
Scientific Reports on 26 October 2020,
Peter Kloess of the
Department of Integrative Biology and
Museum of Paleontology at the
University of California, Berkeley,
Ashley Poust, also of the Department of Integrative Biology and Museum of Paleontology at the University of California, Berkeley, and of the
Department of Paleontology at the
San Diego Natural History Museum, and
Thomas Stidham of the
Institute of Vertebrate Paleontology and Paleoanthropology of the
Chinese Academy of Sciences, the Center for Excellence in Life and Paleoenvironment, and the
University of Chinese Academy of Sciences, add to this assemblage by describing a more than 12 cm long pelagornithid dentary fragment, University of California Museum of Paleontology (UCMP) 323792.

(a) Map of Antarctica with location of Seymour Island highlighted. (b) Generalized geologic map of Seymour Island. Numbers indicate localities containing Antarctic Pelagornithid specimens: (1) UCMP RV8405; (2) DPV 13/84; (3) Specimen MLP 78-X-26-1; (4) IAA 1/95; (5) IAA 1/906; (6) UCMP RV870; and (7) Specimen USNM 494035. Kloess et al. (2020).
Kloess et al. also revise the stratigraphic placement of the previously reported tarsometatarsus specimen, UCMP 322176, within the La Meseta Formation Pelagornithid assemblage. With these two specimens, the known Pelagornithid record from Seymour Island is now represented by six specimens representing multiple taxa; three from the early Eocene (including the presence of a giant-bodied specimen in a temporal period previously represented only by large-sized individuals) and three from the middle to late Eocene (represented by giant-sized individuals). A nearly complete Pelagornithid humerus awaits formal description and would add to this collection of specimens.
The geology of Seymour Island records Cretaceous to latest Eocene marine strata along with Pliocene to Pleistocene glaciomarine deposits restricted to the northern portion of the island. These Eocene strata contain deltaic, estuarine, and shallow marine deposits filling an incised valley, and the Eocene sediments have been subdivided using two different approaches. The first method designates 'Telm' units (an acronym for Tertiary Eocene La Meseta) based on lithofacies, and the second relies on unconformities to divide the La Meseta Formation into allomembers. Some authors have gone further in elevating the uppermost Submeseta Allomember to formation status and subdividing that new formation into the Laminate, Turritella, and Superior Allomembers (formerly the Submeseta I, II, and III units, respectively). Kloess et al. use both the Telm units and the allomembers in accordance with the work of various authors, and refer the reader to the combined stratigraphic column for correlations.

Geochronological context and estimated body size classes of known Pelagornithid specimens from Antarctica. Left, stratigraphic section of the La Meseta and Submeseta Formations. and pelagornithid specimens recovered from Seymour Island. Furthest right, the pelagornithid specimen found in East Antarctica (USNM 494035). Two methods (middle columns) are shown for the subdivision of the La Meseta and Submeseta strata. Numbers and pelagornithid outlines correspond to specimen localities displayed above. The colors and relative sizes of Pelagornithid outlines correspond to the 'large' (in gray) and 'giant' (in black) size-types. Kloess et al. (2020).
Recently, there has been uncertainty regarding the age of the La Meseta and Submeseta Formations. A combination of age-dating methods, such as strontium isotopes, magnetostratigraphy, and lithostratigraphy, indicate that the La Meseta Formation is early to middle Eocene in age and the Submeseta Formation is middle to late Eocene in age. However, studies of Dinoflagellate cysts support a middle to late Eocene age for the La Meseta Formation. Kloes et al. refer to the dates generated from the combined methodology because of the inclusion of the Submeseta Formation into these geochronological and stratigraphic studies.
The La Meseta and Submeseta Formations have yielded many Avian fossils, including Penguins, Falconiforms, Procellariiforms, Anseriforms, Palaeognaths, and Pelagornithids. The Pelagornithids from these deposits are represented by six published specimens. One partial maxilla: MLP 08-XI-30-42 was recovered from locality DPV 13/84. This locality has been assigned to multiple, parallel subunits of the La Meseta Formation: within Telm 7; at the base of level 38; and within the Turritella Allomember (equivalent to the Submeseta II Allomember. The Turritella Allomember has been dated between 41.1 and 37.7 million years on Seymour Island.
Another maxillary fragment (MLP 78-X-26-1) was described first by
Eduado Tonni and Alberto Cione. These authors placed the specimen within the upper portion of the La Mesesta Formation, though its location was not shown on their included maps. A published catalogue of La Meseta fossil material housed in the Museo de La Plata confirms that the specimen comes from an unknown locality and the authors attribute it to the highest stratigraphic unit, Telm 7. MLP 78-X-26-1 was later assigned to locality DPV 13/84 within the Submeseta II Allomember but it is unclear how the assignment to this locality was determined. We use the conservative placement of MLP 78-X-26-1 within Telm 7, which encompasses the Turritella and Superior Allomembers (formerly Submeseta II and III, respectively), to indicate this specimen’s stratigraphic placement. Telm 7 has been dated between 41.1 and 34.0 million years old.
A distal Pelagornithid humerus (MLP 12-I-20-4) was recovered from locality IAA 1/95. This Pelagornithid-bearing locality is in the same stratigraphic horizon as the Mammal-bearing locality IAA 1/90, located within a Naticid Gastropod-dominated conglomerate layer in Telm 5 and the
Cucullaea I Allomember. Additionally, a recently published rostral dentary fragment (IAA-PV 175) from locality IAA 1/90 was
described by
Carolina Acosta Hospitaleche and Marcelo Reguero confirming that identification and publishing the first image of this specimen (however, without identifying its specimen number). In a review of the Phorusrhacid material from the Cretaceous and Palaeogene of Antarctica, Marcos Cenizo reassigned the tarsometatarsus (identifying the specimen number as 'UCR 22176') to the Pelagornithidae. Claudia Tambussi and Federico Degrange incorrectly refer to this specimen as cast UCR 22175; the specimen number used (revised as UCMP 322175), corresponds to a fragment of a premaxilla reassigned from Phorusrhacidae to an unknown genus and species of Palaeognath.
UCMP 322176 has been attributed to the Submeseta Formation. However, a review of the original locality information indicates that rather than the Submeseta Formation, the tarsometatarsus was recovered from the same stratigraphic horizon as UCMP locality RV8200 (formerly RV-8200) within Telm 5 (the La Meseta Formation). UCMP locality RV8200 is equivalent to the locality DPV 6/84, stratigraphically higher than IAA 1/90, and within the Cucullaea I Allomember. The Cucullaea I Allomember is dated between 51.6 and 49.1 million years, and Telm 5 is dated between 51.3 and 46.2 million years. These dates yield an estimate of 51.3–49.1 million years for the age of fossils from UCMP locality RV8702.
For the first time, Kloess et al. include high resolution images of the original tarsometatarsus fossil to highlight aspects of its morphology because previous authors have published only images of casts made from UCMP 322176. A detailed description of this tarsometatarsus, including observations of characters that this specimen shares with the Dasornis and Pelagornis morphotypes and size comparison to other known distal tarsometatarsus fragments, was presented by Marcos Cenizo, and the specimen was assigned further to a 'giant' size-type (estimated 5–6 m wingspan). The reassessed stratigraphic placement of this specimen to the La Meseta Formation, where it joins other La Meseta specimens identified as 'large' size-types (estimated 3.5–4.5 m wingspan), indicates that Seymour Island was inhabited by two different size classes of Pelagornithids during the early Eocene.

Photographs of the original fossil material of the pelagornithid distal right tarsometatarsus UCMP 322176 in: dorsal (a), medial (b), plantar (c), lateral (d), and distal views (e). Residual mold lines from the castmaking process can be observed on the medial, lateral, and trochlear surfaces (b), (d), and (e). Osteological abbreviations: cid canalis interosseus distalis, dlr dorsal longitudinal ridges, flc fovea ligamentum collateralium, fm I fossa metatarsi I, fvd foramen vasculare distale, pmp processus medianoplantaris, pp plantar 'pit', tr metatarsal trochlea. Kloess et al. (2020).
UCMP 323792 is a partial left dentary 12.2 cm long, preserving a region of the mandible rostral to the intraramal joint. The dentary is relatively flat with a prominent mandibular groove (lateral longitudinal sulcus, external groove, neurovascular sulcus, or longitudinal furrow) tracing the ventral length of the specimen. The groove is asymmetric, with a flatter ventral side compared to the gently sloping dorsal side. At the caudal end of the fragment, the groove is 11.4 mm dorsal to the ventral edge of the dentary, and 9.0 mm dorsal to that edge at the rostral end. The groove is 19.5 mm ventral to the dorsal edge of the dentary at the caudal end, and 15.8 mm ventral to the dorsal edge of the rostral preserved end. Overall, the groove approaches the ventral edge of the bone rostrally.
Left
Pelagornithid dentary fragment UCMP 323792 in lateral (a), medial (b),
and dorsal views (c). (d) Line drawing of dorsal view to elucidate
location of pseudoteeth. Pseudoteeth depicted as dashed outlines. Note
on PT1 the presence of a mediocaudal crest and its tip is offset from
the midline. Abbreviations: MC mediocaudal, PT pseudotooth. Kloess et al. (2020).
Though the rostroventral portion of the dentary is missing, the dorsal edge of the mandibular groove and Meckel’s canal are visible. This specimen shows that the groove is immediately adjacent to Meckel’s canal and thus likely related to the nutrient supply for the growing multipart rhamphotheca.
Given that UCMP 323792 derives from an inexact location within the dentary rostral to the intraramal joint, Kloess et al. compared measurements of its dorsoventral height to those collected from published images of complete dentaries from the largest Pelagornithids, Pelagornis chilensis, and Pelagornis sandersi, both 'giant' size-type Pelagornithids from the Miocene and late Oligocene, respectively. Measurements from these specimens were collected from the base of the most rostral and most caudal PT1s, as well as the PT1 closest to the midpoint between them. The rostral height measurement from UCMP 323792 (24.8 mm) is greater than the most rostral heights of Pelagornis chilensis, and Pelagornis sandersi, 19.9 and 9.0 mm respectively, and just less than heights from their midpoints, 28.7 and 25.5 mm respectively. The most caudal heights of these species (Pelagornis chilensis: 40.1 mm; Pelagornis sandersi: 34.2 mm) is greater than the caudal height of UCMP 323792 (30.9 mm). Based on these measurements, UCMP 323792 falls well within the range of heights of these largest known Pelagornithids and the fragment likely comes from near the rostrocaudal midpoint of the dentary.
While UCMP 323792 has low, worn pseudoteeth similar to the oldest, smallest, and geographically closest Pelagornithid to Antarctica, Protodontopteryx ruthae, the preserved pseudoteeth in the maxilla and mandible of Protodontopteryx ruthae from the early Paleocene of New Zealand are weathered to the point that size class identification is impossible though it can be noted that the specimen exhibits approximately regular spacing of its pseudoteeth. The entire length of the preserved right dentary of Protodontopteryx ruthae measures less than the preserved length of UCMP 323792, and helps to document the very large diversity of body sizes within the clade.
A wide shallow groove is visible along the ventral dentary. Gerald Mayr and David Rubilar-Rogers used the term 'neurovascular sulcus' for both the medial and lateral mandibular grooves in their figures of Pelagornis chilensis. The portion of the dentary dorsal to the groove is convex in profile and dorsoventrally taller (16.4 mm rostrally and 24.9 mm caudally) relative to the ventral dentary edge (about 6.3 mm). Evidence of erosive events (pits, black Lichen, and weathering cracks) are readily visible on this side of the specimen. Similar evidence of wear can be observed on the rounded and broken pseudoteeth. Lichen and related pits, resulting from apothecia, have been observed on fossils from the Submeseta Formation and their presence indicates the dentary had been exposed medial side up at the subaerial surface prior to its discovery.
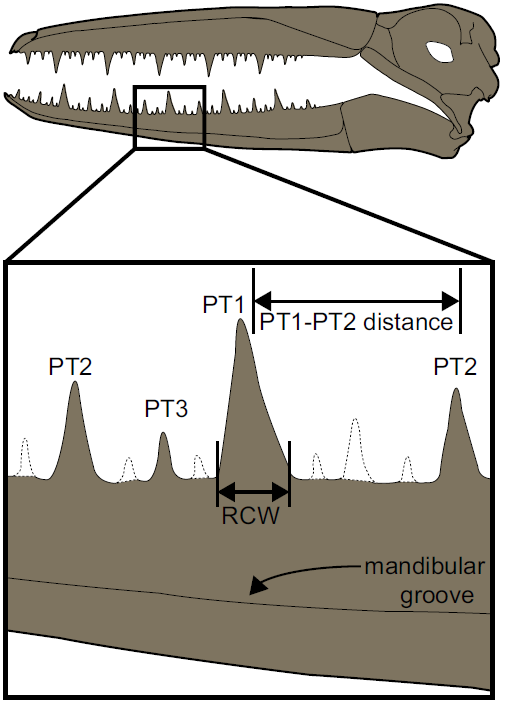
Four low, worn pseudoteeth are visible. All of the pseudoteeth exhibit the remnants of a mediocaudal ridge, similar to those observed in other Pelagornithids. Based on the regular pattern of pseudotooth spacing observed in other pelagornithids, some of the smaller bony projections may have been worn away from UCMP 323792, and the possibility that even smaller, intermediately-spaced pseudoteeth (i.e. PT4s and PT5s) were present previously and also worn away cannot be ruled out. It is likely that there is a pseudotooth missing between the PT1 and the caudalmost PT2, and one missing rostral to the most rostral PT2.

Diagram approximating the location of the dentary fragment, UCMP 323792, within a Pelagornithid mandible. PT numbers indicate size class of pseudotooth, with PT1 representing the largest 'teeth' and PT3 the smallest 'teeth'. Bars indicate method for measuring rostrocaudal width and the distance between PT1 and PT2s. Dashed lines indicate the possible locations of missing pseudoteeth. PT pseudotooth, rostrocaudal width. Kloess et al. (2020).
The distance measured from the existing PT1 to each of the remaining PT2s is greater than similar measurements from the largest Pelagornithids, Pelagornis chilensis, and Pelagornis sandersi. Since there is only one PT1 present in UCMP 323792, Kloess et al. measured the distance between the PT2s present as a proxy for calculating the space intervening between PT1s, and estimate that distance as 66.5 mm. This estimate of pseudoteeth spacing from UCMP 323792 is greater than all measurements compiled from various Pelagornithids, by Antoine Louchart, Vivian de Buffrénil, Estelle Bourdon, Maïtena Dumont, Laurent Viriot, Jean-Yves Sire, except those within the larger range of measurements from Pelagornis chilensis (ranging from 53.9 to 76.0 mm). The base width of the PT1 present (14.7 mm) also is larger than those compiled by Louchart et al. except for the larger measurements from Pelagornis chilensis (12.5–15.6 mm) and Pelagornis longirostris (13.3–15.9 mm), a Pelagornithid of unknown Cainozoic age with cranial dimensions similar to Pelagornis chilensis. Based on these measurements, UCMP 323792 may represent one of the largest Pelagornithids found to date.
The first reconstruction of a Pelagornithid rostrum from Dasornis (Odontopteryx) toliapica indicates a repeating pattern of pseudoteeth. Hildegard Howard's description of Pelagornis (Osteodontornis) orri provided details of the pattern of spacing and placement of pseudoteeth sizes; the largest pseudoteeth were spaced regularly along the length of the rostrum with the interstitial space bisected by moderately-sized pseudoteeth and then further split evenly by the presence of the smallest pseudoteeth. Although this pattern of pseudoteeth spacing has been observed in Pelagornithid specimens identified since Howard’s observations, exceptions have been noted, for example at the tip of the rostrum or duplicate PT2s between PT1s.
Louchart
et al.
reported known differences in the distribution and size of pseudoteeth between Odontopterygiform species and calculated a regression line, based on the space between the largest pseudoteeth (PT1s) and a ratio of the rostrocaudal widths of pseudoteeth of different sizes, that supports a proposed mechanism for pseudotooth size and spacing based on inhibition zones. UCMP 323792 does not follow the pattern described by Louchart
et al., but rather maintains a relatively low (extrapolated) intervening space between PT1s, as well as a lower value for the ratio of pseudotooth widths. To account for the difference between the reported regression line To account for the difference between the reported regression line and calculations from UCMP 323792, Kloess
et al. consider taphonomic effects which have altered the specimen including, but not limited to, weathering of the pseudoteeth diminishing in vivo base widths and the loss of PT1s. Alternatively, measurements from UCMP 323792 may not align with the regression line for biological reasons, such as: this specimen may reflect a different ontogenetic stage or clade of Pelagornithid from those specimens used in previous calculations, and thus may have a different pattern of pseudotooth spacing altogether. However, the regression line as published does not prescribe identification to taxon or ontogenetic age based on Pseudotooth measurements and would require the addition of more specimens to properly make these assessments.
The new pelagornithid dentary fragment described above (UCMP 323792) provides an opportunity to reexamine the previously identified Antarctic Pelagornithid fossils, offering insight into the presence of coexisting morphotypes in the Southern Ocean throughout the Eocene. The measured and extrapolated spacing between pseudoteeth of UCMP 323792 point to a giant individual, but the use of widths from worn pseudoteeth may not be well-suited for detailed comparisons. Most rostral specimens of Pelagornithids are partial, fragmentary, or taphonomically altered. If the metric of Louchart et al. is inappropriate for incomplete specimens, such as UCMP 323792, the general utility of many specimens for size regression is called into question, as is the use of spacing for taxonomic discrimination except in cases where intact segments can be confidently positioned within the oral cavity. The regression calculated by Louchart et al. also excluded specimens with the smallest pseudoteeth (PT4s and PT5s), those most susceptible to erosion and weathering, and thus potentially excluded exceptionally preserved specimens.
The updated stratigraphic context for the Pelagornithid distal right tarsometatarsus (UCMP 322176) implies the presence of a 'giant' Pelagornithid taxon in the early Eocene of Antarctica. The other Pelagornithid material from the early Eocene La Meseta Formation (a distal humerus, MLP 12-I-20-4, and a partial dentary fragment, IAA-PV 175) exhibit morphology and size similar to 'large; Pelagornithids, such as cf. Gigantornis sp. The stratigraphically-reassigned tarsometatarsus (UCMP 322176) possesses morphological affinities to both the Dasornis and Pelagornis morphotypes though its width is greater than tarsometatarsi identified as 'giant' Pelagornis taxa. The difference in sizes of these specimens suggests that 'large-' and 'giant-sized' Pelagornithid taxa co-occurred in the early Eocene of Antarctica, and that the giant size class of Pelagornithids evolved quite early in their history.
With the reassignment of a recently published partial dentary (IAA-PV 823) as a Perciform Fish, the late Eocene of Seymour Island is currently unambiguously represented by only 'giant' size-type specimens of Pelagornithids. However, the fragmentary humerus (USNM 494035) from Mount Discovery indicates the presence of multiple Pelagornithid size-types and taxa across Antarctica during this time. Therefore, it would appear that the two Pelagornithid morphotypes, and the largest two Pelagornithid size-types spanned from the early Eocene to the late Eocene of Antarctica. The Eocene La Meseta and Submeseta Pelagornithid specimens suggest the presence of an unnamed species larger than known Eocene taxa. Known specimens that approach the size of these Antarctic specimens have been recovered from Oligocene and Miocene strata, but not yet the Eocene. Furthermore, the reassigned La Meseta tarsometatarsus (UCMP 322176), with characteristics intermediate between the two accepted morphotypes, may represent an unnamed species larger than all known Pelagornithid taxa. None of the specimens from Antarctica have been identified to genus, nor have any of them been used to establish new taxonomic names. However, there are likely at least two taxa (or species lineages) present through the Eocene of Seymour Island, and that only with the discovery and description of more overlapping skeletal elements may we begin to evaluate the alpha level diversity of Pelagornithids present in this ancient ecosystem. Nevertheless, these unnamed remains are a tantalizing suggestion that the largest Bird that ever flew may have soared its way over the Antarctic seas during an Eocene with a unique, and distinctively large-bodied, coastal Avifauna. In addition, the distribution of Pelagornithid body sizes in the same pelagic Antarctic ecosystem likely reflects ecological differences related to diet or foraging strategy, and indicates stability in those ecological niches through much of the Eocene. This updated fossil record of Pelagornithids on Seymour Island reinforces the ideas that along with Penguins and Palaeognaths, Pelagornithids were a common and even a dominant avian clade throughout the Eocene of Antarctica, and potentially competed with other soaring Birds for foraging and nesting spaces. These Pelagornithids would have occupied a high trophic level in Antarctic seas, a role today filled by Albatrosses and other pelagic Avian clades, and the combined utilisation of marine resources by pelagic Birds and Penguins seen today likely extended into the deep past.
See also...



Online courses in Palaeontology.
Follow Sciency Thoughts on Facebook.
Follow Sciency Thoughts on Twitter.