The end-Triassic Mass Extinction is one of the largest known biological crises of the Phanerozoic and is regarded as one of the 'Big Five'. This extinction has been linked with voluminous volcanism during the emplacement of Central Atlantic Magmatic Province and its associated environmental effects. These effects include global warming and ocean anoxia. Existing evidence suggests that basinal marine anoxia was widespread on the northern Panthalassan margin of Pangaea and that intense shelf euxinia also became widespread in the latest Triassic–earliest Jurassic of Western Europe, but some of these conditions developed, some roughly150 thousand years after the onset of the end-Triassic Mass Extinction. Additional findings from seawater uranium isotopes in the Lombardy basin of western Tethys suggest an increase in the extent of anoxic deposition through the Triassic-Jurassic boundary. However, in other oceans, clear evidence for widespread anoxia in the latest Rhaetian that directly coincides with the beginning of end-Triassic Mass Extinction has not been recorded, leaving its role as the cause of the marine component of the end-Triassic Mass Extinction questionable.
Carbonate-associated sulphate in bulk marine carbonate and biogenic calcite is widely used to reconstruct the primary seawater sulphate sulphur isotope composition during major redox perturbations of the Earth surface system. The proportion of sulphur³⁴ in seawater is dynamically controlled by variations in the fluxes and isotopic compositions of riverine sulfate sources and marine pyrite burial. The removal of sulfate from the oceans via gypsum precipitation does not impart an isotopic fractionation, but this removal makes the global sulfate reservoir smaller and, therefore, more isotopically susceptible to changes in other fluxes. The production and burial of pyrite represent a primary redox-sensitive pathway in the marine sulphur cycle, which drives a large offset between the sulphur isotopic composition of the seawater sulphate and sedimentary pyrite pools, and thus may control variations in the sulphur isotope composition of oceanic sulphate through time. Large and rapid global-scale sulphur isotope perturbations, as well as the small ocean sulphate reservoirs needed to produce them, seem to be a feature of major deoxygenation events of the Phanerozoic. Although there is some evidence in the sedimentary pyrite isotope record that suggests the regional development of marine anoxia at the end-Triassic Mass Extinction, direct records of changes in the marine sulphate pool and therefore impacts on the global sulphur cycle are undocumented.
In a paper published in the journal Science Advances on 9 September 2020, Tianchen He of the School of Earth and Environment at the University of Leeds, Jacopo Dal Corso, also of the School of Earth and Environment at the University of Leeds, and of the State Key Laboratory of Biogeology and Environmental Geology at the China University of Geosciences, Robert Newton, Paul Wignall, and Benjamin Mills, again of the School of Earth and Environment at the University of Leeds, Simona Todaro and Pietro Di Stefano of the Department of Earth and Marine Sciences at the University of Palermo, Emily Turner and Robert Jamieson, again of the School of Earth and Environment at the University of Leeds, Vincenzo Randazzo, also of the Department of Earth and Marine Sciences at the University of Palermo, Manuel Rigo of the Department of Geosciences at the University of Padova, Rosemary Jones of the Department of Earth Sciences at the University of Oxford, and Alexander Dunhill, once again of the School of Earth and Environment at the University of Leeds, report three open marine sulphur isotope composition of oceanic sulphate profiles from Sicily (Mount Sparagio Section), Northern Ireland (Cloghan Point Section), and British Columbia (Black Bear Ridge Section). These derive from both Tethyan and Panthalassan locations; the first two
sections archive well-preserved, shallow-water, peritidal, micritic,
and shelly limestones and shell materials; and the last section consists
of open-shelf, organic-rich, and Bivalve-rich marly limestone. The
sections span the Norian to lower Hettangian and record the major losses
of the end-Triassic Mass Extinction. Therefore, they provide a window
into the possible links between the ecosystem response and marine redox
variations in Late Triassic oceans over a broad area.

Simplified paleogeographical map for Triassic-Jurassic transition showing localities for all three studied sections. Yellow filled triangles indicate the location of studied sections. He et al. (2020).
All sulphur isotope composition of oceanic sulphate profiles from three different localities show similar trends, although the absolute values vary between the European and North American sections. In all sections, a large positive sulphur³⁴ isotope shift with a magnitude of more than 10 per mil (‰) is seen in the latest Rhaetian (roughly 201.5 million years ago) and coincides precisely with the extinction horizon. Two consecutive positive sulphur³⁴ excursions are shown at the Mount Sparagio Section, while only a single spike is seen at the other two sections. At the Cloghan Point Section, only the falling limb of the positive excursion was recovered because of the absence of suitable bulk carbonate or shell material below this level. The pre- and postexcursion baseline values for the two Tethyan sections are between 15 and 20‰, which are close to the existing global sulphur isotope composition of oceanic sulphate and evaporite dataset for the Late Triassic. By contrast, the sulphur isotope composition of oceanic sulphate record at the Panthalassa Black Bear Ridge Section generally yields more positive baseline values and a slightly larger positive swing. This is likely due to the development of sulphate isotopic and concentration heterogeneity between Tethyan and Panthalassan sites under low sulfate conditions. He et al. note also that the positive sulphate sulphur³⁴ excursion at the Black Bear Ridge Section is mirrored by synchronous positive sulphur³⁴ shifts in sedimentary pyrite at a deeper site (Kennecott Point section) in eastern Panthalassa, suggesting a coupled behavior in both marine oxidised and reduced sulphur sinks.
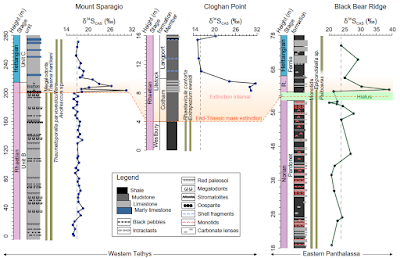
Sulphur isotope composition of oceanic sulphate profiles from Late Triassic to Early Jurassic for the three studied sites of the Tethys and Panthalassa oceans. R., Rhaetian. The orange shadowed field indicates the extended extinction interval following the major mass extinction horizon. The light green field indicates a hiatus between Norian and Rhaetian at the Black Bear Ridge Section. Dark green bars represent the fossil occurrence ranges. Vertical dash lines indicate pre- or postexcursion average baseline values. He et al. (2020).
He et al. calculated the age model at the most stratigraphically complete Tethyan Mount Sparagio Section. The duration of the shift from the baseline value (about 16 to 17‰) to the first peak value (roughly 31‰) is estimated to take about 50 000 years with the assumption of a constant sedimentation rate and a Rhaetian duration of 4.1 million years. This time frame is broadly in agreement with the equally short-lived major phase of the extinction, which was proposed to last for about 40 00 years. Thus, the observed sulphur³⁴-positive excursion event in the latest Triassic appears to represent an extreme and short-lived perturbation when compared to other similar positive sulphur³⁴ isotope events during, for example, the end-Permian Extinction (roughly 100 000 years), Toarcian Oceanic Anoxic Event (roughly one million years), and Cretaceous Oceanic Anoxic Event (roughlu 500 000 years).
The observed positive swing in the sulphur isotope composition of seawater sulphate in the latest Triassic could have been driven by an increase in the net burial of sedimentary pyrite under expanded anoxic/euxinic conditions. These conditions result in enhanced microbial sulphate reduction, leading to an enhanced pyrite burial flux on the continental shelves and slopes when there is sufficient supply of available iron and organic matter. Because pyrite is depleted in the heavier isotope sulphur³⁴, elevated burial fluxes on a global scale would drive the seawater sulphate sulphur³⁴ to more positive values. The oxidative biotic pathway of the global sulpjur cycle may also have the potential to drive seawater sulfate sulphur³⁴ enrichment to some extent via microbial sulphide oxidation by some sulphide-oxidising microorganisms. However, the contribution of this oxidative metabolic pathway to the oceanic sulfate pool remains unclear, and there is no obvious mechanism for it to have driven a prolonged positive sulphur isotope excursion in the global seawater sulphate inventory. On a larger scale, it may be possible to drive sulphur isotope variations by altering the weathering rates of continental pyrite and gypsum; here, a geologically sudden increase in seawater sulphur³⁴ might represent a cessation of pyrite weathering and a switch to an isotopically heavy riverine flux.
To investigate the response of seawater sulphate sulphur³⁴ to the variations of oceanic sulphate inventory and the degree of change in the net pyrite burial flux, He et al. applied a time-dependent sulphur cycle single-box model. The model assumes that the isotopic composition of the pyrite and gypsum weathering fluxes remain constant, and experiments then alter the pyrite input and output fluxes through either weathering or burial. In the model, a substantial increase in pyrite burial by approximately a factor of 5 and a very small marine sulphate reservoir (less than 1 mM) is required to replicate the magnitude and timing of the sulphur isotope composition of oceanic sulphate shift. This version of the model fixes the isotopic enrichment of buried pyrite at 30‰ more negative than contemporaneous seawater sulphate, but the expansion of euxinia may have increased this enrichment factor; thus, He et al. also experiment with a scenario in which this is increased to 40‰ during the event. This experiment has a very similar requirement for a large increase in pyrite burial and very low seawater sulphate concentration. Note that it is the size, direction, and duration of change that are the important foundations of our modeling approach. Differences in regional sulphate isotope baselines have no impact on the conclusions from the modeling work, as a similar sized isotope excursion is present in all records. Replicating the change in sulphur³⁴ by reducing pyrite weathering rates while maintaining the same gypsum weathering flux is much more difficult and requires a complete cessation of pyrite weathering and extremely low ocean sulphate (about 0.1 mM). Even then, the shape of the excursion is not readily reproducible, as the very low sulphate concentrations mean that the system rapidly recovers from the perturbation.

Sulphur cycle box model outputs. (A) and (B) Increased in the pyrite burial rate under different values for the starting oceanic sulfate inventory, with tests of 1 mM (A) (yellow) and 0.33 mM (B) (red). For both scenarios, a step increase in pyrite burial is assumed to occur at t = 0 over a period of 50 ka, which represents the end-Triassic Mass Extinction. Both models assume the same increase in pyrite burial rates, which ranges from 2- to 10-fold to create the shaded area, with the centerline showing a fivefold increase. The best fit to the data occurs for marine sulphate concentration (SO₄) = 0.33 mM (B). (C) Attempts to fit the sulphur isotope composition of oceanic sulphate profiles data by instead reducing the pyrite weathering rate to zero over the same 50 000 year time frame. Here, regardless of (SO₄), the shape of the curve cannot be fit. This is because creating the large excursion this way requires extremely low (SO₄), and, in these circumstances, the system is quick to regain isotopic stability. (D) to (F) Repetition of these experiments with the addition of a change in the enrichment factor sulphur³⁴ between oceanic sulfate and sedimentary pyrite and continuation to produce a better fit when (SO₄) = 0.33 mM. He et al. (2020).
The maximum marine sulphate concentrations can be independently estimated using the maximum rate of change in sulphur isotope composition of oceanic sulphate. The 'rate method' model gives an upper estimate for marine sulphate of about 0.2 to 1.1 mM for the interval through the Late Triassic–positive isotope excursion event. The lower end of these maximum estimates is consistent with the calculations inferred from our sulfur cycle box model. Therefore, the intervals predating and during the positive sulphur isotope excursion event appear to be characterised by a scarcity of oceanic sulphate when compared to a higher fluid inclusion–based estimate of at least 13 mM during the Carnian, although this was about 20 million years earlier. The development of a low sulphate ocean in the later Triassic was likely caused by substantial evaporite deposition. As shown in global compilations for this interval, minimum estimates of global halite deposition suggest a 16-fold increase from the Middle to Late Triassic. By contrast, the earlier part of the Triassic experienced a low level of evaporite occurrence following the end-Permian extinction. Late Triassic evaporites were deposited in newly formed rift basins that developed in an arid climate as Pangaea began to break up. When examined on a regional scale, for example, evaporite deposition became widespread surrounding the North Atlantic rift (northeastern Grand Banks, Oranian meseta, and Western Europe) during the Late Triassic and subsequently peaked in the Earliest Jurassic.
He et al.'s finding of low marine sulphate concentrations preceding an episode of massive pyrite burial in the latest Triassic adds to an increasing number of studies that link low seawater sulphate with the expansion of anoxic waters in the oceans. He et al. propose a conceptual model to link these observations. Marine sulphate and organic carbon availability exert a major control over the balance between three microbially mediated biogeochemical pathways in marine sediments: Microbial sulphate reduction (sulphate + formaldehyde → hydrogen sulphite + bicarbonate), methanogenesis (acetate + hydrogen → methane + carbon dioxide and carbon dioxide + hydrogen → methane + water), and the anaerobic oxidation of methane (methane + sulphate → bicarbonate + hydrogen sulphite + water). Under high sulphate conditions such as the modern ocean, microbial sulphate reduction consumes large amounts of organic carbon, while methane is produced deeper in the sediment where sulphate has been depleted. The overlying sulphate-rich pore water fuels anaerobic oxidation of methane and prevents substantial benthic methane escape, therefore limiting bottom-water oxygen consumption. In contrast, under conditions of low sulphate availability, the balance of processes oxidising organic matter in marine sediments shifts in favor of methanogenesis, as occurs widely in freshwater sediments (e.g. lakes), where sulphate supply is usually limited. Lower sulfate concentrations bring the sulphate-methane transition zone closer to the sediment-water interface and reduce the amount of organic matter consumed by microbial sulphate reduction, ultimately increasing the organic carbon flux to methanogens and limiting the capacity for anaerobic oxidation of the resulting methane. The organic matter reaching the zone of maximum methanogenesis will also have increased reactivity. The result is a greater flux of methane from the sediment, leading to increased aerobic respiration of methane close to the sediment-water interface placing an increased burden on bottom-water oxygen levels.
In the modern system, around 98% of all buried organic carbon in the ocean is stored in continental margin sediments. On average, around 20% of the global organic carbon flux (roughly 191 Tmolof carbon per year) to the seafloor is processed via microbial sulphate reduction, and about 3 to 4% is converted to methane, giving an annual methane flux from seafloor of about 5.7 to 7.6 Tmol of methane per year. He et al. calculate that a drawdown in oceanic sulphate concentration by roughly 97% from 29 mM (modern value) to 1 mM will reduce the rate of microbial sulphate reduction by a similar amount and that the excess organic matter will all be used by methanogens (i.e. they now process about 22 to 23% of the organic carbon), then the methane flux would rise to around 42 to 44 Tmol methane per year. This calculation is conservative, since it does not take into account any increase in reactivity of the organic matter reaching the methanogenic zone. Furthermore, suppression of anaerobic oxidation of methane under these low sulphate conditions would make it easier for this methane to reach the water column and consume free oxygen. Making more detailed calculations on the expected impact of low sulfate conditions on water column oxygen demand requires further modeling, which is beyond the scope of He et at.'s study, but their calculations demonstrate that there is clear potential for at least a six- or sevenfold elevation in the methane flux at the sediment-water interface and a concomitant increase in the global consumption of benthic oxygen. Note that these elevated demands on bottom-water oxygen exist where sulphate concentrations are low and before any additional drivers from the release of volcanic carbon dioxide.
Finding evidence for elevated aerobic methane oxidation under low sulphate conditions in the sedimentary record is not simple because the resulting dissolved inorganic carbon flux, while large when considered in the context of dissolved oxygen uptake, is small compared to the abundance of ocean dissolved inorganic carbon, especially when oxidation takes place in the water column as proposed. Isotopically depleted carbonate cements form from pore waters and are a common feature of the sedimentary record and so do not provide definitive evidence. Calcifying organisms living at the sediment-water interface are likely to provide the best archive for recording this process, evidence for which has been recognized in high-latitude late Cretaceous Bivalves,
A key feature of our conceptual model is that sulphate poor conditions are established before volcanic perturbation, likely by widespread evaporite deposition. Previously, authors have explained the link between the expansion of marine anoxia during large igneous province-driven warming and extinction events via the decreased solubility of oxygen in warmer waters and increased productivity and oxygen demand driven by increased weathering fluxes of nutrients from land and the recycling of phosphorus once euxinic water column conditions are established. The higher bottom-water oxygen demand of a steady-state Earth system with a small marine sulphate reservoir will predispose the oceans to the rapid expansion of anoxic conditions via these mechanisms. In addition, a low sulphate ocean is likely to impose some additional feedbacks once warming has been initiated: The rate of methanogenesis is highly temperature sensitive, so methane production will increase with sediment temperature, a situation amplified by the reduced depth to the methanogenic zone under low sulphate conditions. Increased marine organic matter production will increase the delivery of organic matter and its reactivity to the methanogenic zone in sediments, again adding to increased methane fluxes across the sediment-water interface and oxygen consumption from methane oxidation. Pyrite burial will increase as anoxic conditions expand, creating downward pressure on marine sulphate concentrations, although this may be countered by bigger fluxes of weathered sulphate from land. Elevated global marine methane production may also promote methane release to the atmosphere and thereby contribute to warming trends initiated by the large-scale release of volcanic carbon dioxide, although much of the additional methane production is likely to be oxidised in the water column. These additional feedbacks may explain why the expansion of anoxic conditions is more severe under low sulphate conditions and why not all large igneous province-driven warming events create widespread oxygen depletion.

Conceptual model of the methane-oxygen link under high and low sulphate conditions. (A) The fate of organic carbon in the modern high sulphate ocean: More organic carbon and methane are oxidised by sulphate with negligible benthic methane flux, which limits water column oxygen demand. (B) The effect of enhanced methanogenesis in a low sulphate setting: The proportion of organic carbon available for methane production is increased, sulfate-driven anaerobic oxidation of methane is suppressed, and methane production moves closer to the sediment surface producing a high benthic oxygen demand. Red arrows in (B) indicate acceleration of biogeochemical pathways relative to modern, whereas dotted black arrows indicate retardation. (C) and (D) The envisaged oxygen depletion responses of the ocean to the same carbon dioxide forcing under high and low sulfate conditions. Sulphate is thought to be removed by evaporite deposition. Marine anoxia is exacerbated by the increased oxygen demand as net seafloor methane fluxes increase during warming. MG, methanogenesis. SMTZ, sulfate-methane transition zone. He et al. (2020).
Although anoxia may not have developed on the deep ocean floor during the Triassic-Jurassic transition, other geochemical evidence, in the form of enrichment of redox-sensitive elements (e.g. maganese and molybdenum) and nitrogen isotope fluctuations, suggests that there was a major intensification of the mid-water oxygen minimum zone in the Panthalassa Ocean at the time. Tangible evidence for this is seen where the oxygen minimum zone impinged on the western margin of the Pangean supercontinent, leading to extensive black shale deposition in Western Canada. Euxinia also became extensive in the latest Triassic shelf seas of Western Europe, both during and at the termination of the mass extinction phase. Uranium isotope data from marine carbonates provide a possible measure of ocean redox conditions with negative excursions of the proportion of uranium²³⁸ values signifying enhanced reduction from uranium (VI) to uranium (IV). Such a signal, seen at the start of the mass extinction, suggests a major increase in the area of anoxic deposition that lasted for about 50 000 years.
He et al.'s sulphur isotope composition of oceanic sulphate excursions reveal a similar link between the onset of mass extinction and an anoxia-driven isotopic excursion. The link is most clearly seen in western Tethys where Megalodont Bivalves and the Foraminifer Triasina hantkeni are suddenly lost at the onset of the positive shift. Although there is no direct evidence for anoxia at this peritidal location, some contemporaneous anoxic sedimentary matrices are seen at a neighboring site that was also connected to the western Tethys. There is a hiatus in the Panthalassan section (Black Bear Ridge Section), but the extinction level is still recorded. This occurs in the dysoxic strata of the basal Fernie Formation, where the last Rhaetian Conodonts disappear, and is coincident with the sulphur³⁴ isotope excursion. The extinction of Monotid Bivalves at Black Bear Ridge Section marks an earlier crisis at the end of the Norian, several million years before the end-Triassic event. The end-Triassic extinction is also seen at the Cloghan Point Section, where several Bivalve species, including the Rhaetian marker Rhaetavicula contorta, disappear at the base of the Cotham Member. The lack of limestones at this level precludes measurement of the ratio of sulphur³⁴, but the lowest data point obtained in this section, a short distance above, displays a strongly positive value. In summary, the major sulphur³⁴ isotope excursion found here is best explained by a major pyrite burial event driven by a large-scale, increase in anoxia in the late Rhaetian. He et al.'s age model for the Mount Sparagio Section suggests a 50 000 year duration for the initial positive shift in sulphur³⁴, a time span in remarkable accord with the 50 000 year estimate for the main anoxia intensification during latest Rhaetian based on the contemporary uranium isotope record. Subsequently, the gradual falling limb of the sulphur³⁴ excursion corresponds with the second phase of limited anoxia that extended into the Hettangian. The event also saw the intensification of the Panthalassan mid-water oxygen minimum zone and the deposition of black shales on the Pangean margin and in the shelf seas of Europe. Shallowest water locations, such as the Mount Sparagio Section, remained oxygenated. The coincidence of the sulphur³⁴ excursion with the extinction losses implicates anoxia as an important factor in the crisis.
The late Permian and the Mesozoic Era were punctuated by recurring oceanic anoxic events accompanied by hyperthermal events and enhanced weathering that coincide with the eruption of large igneous provinces. Large positive sulphur isotope shifts in seawater sulphate provide evidence of a greatly reduced marine sulphate reservoir and enhanced pyrite burial for many of these oceanic anoxic events. He et al. explain this generalised coincidence via a mechanistic linkage between low dissolved sulphate, enhanced sedimentary methane generation, and consequent elevated bottom-water oxygen consumption. Hence, He et al. propose that a low sulphate boundary condition before volcanically driven greenhouse warming events makes the expansion of anoxic conditions more likely and that associated feedbacks during the event extend the geographic reach and intensity of anoxia. Many of these events are preceded by increased evaporite burial fluxes, suggesting that this is the mechanism for sulphate removal from the ocean. Hence, the development of widespread anoxia during rapid warming may ultimately trace some of its origins to widespread rifting or other circumstances that create favorable conditions for evaporite deposition.
See also...

Online courses in Palaeontology.
Follow Sciency Thoughts on Facebook.
Follow Sciency Thoughts on Twitter.





































