Geological and geochemical evidence has revealed that the Neoarchean–Paleoproterozoic period was vitally important for Earth’s evolution. The earliest ‘snowball event’ and major glaciation occurred during this period. This was followed by a great oxidation event, which caused an abnormal positive shift in global carbon isotopes and is referred to as the Lomagundi Event. The emergence of oxygen-producing photosynthetic organisms that led to the sudden increase in atmospheric oxygen has been the focus of several studies. However, until now, convincing fossil records from this key geological time interval (i.e. latest Neoarchean to Palaeoproterozoic) are scarce. During this period, the biosphere experienced multiple geological events, but little is known about it and what is known is dependent on molecule clock dating analyses and estimates. To better understand the biosphere during this time, the metamorphosed Palaeoproterozoic deposits of the Hutuo Group at Wutai Mountain in Shanxi Province, China provide an excellent stratigraphic sequence in which to study well-preserved fossil records of this key period in Earth’s evolution.
The Wutai Mountains are located in the Xinzhou area of Shanxi Province, China, between 38°50′ North and 39°05′ North, and between 113°29′ East and 113°44′ East. The Wutai Mountains of the central Trans-North China Orogen are a typical region for the investigation of Precambriansequences. The Precambrian strata in the Wutai Mountains can be divided into the Neoarchean Wutai Group, and the overlying Palaeoproterozoic Hutuo Group, separated by an unconformity.
Geology of Wutai Mountains showing the sample localities. Yin et al. (2020).
The Hutuo Group is distributed in an area of about 1500 km², from northernmost Taihuai-Sijizhuang on the south slope of Wutai Mountain to southernmost Shizui-Dingxiang, and from the upper Taishan River in the east, to Yuanpingqi village in the west. Although the Hutuo Group underwent a major tectonic movement (the ‘Lulianng Movement’) to show strong fold, which has still completely reserved many primary deposited structures, such as cyclothems (alternating stratigraphic sequences of marine and non-marine sediments), wavemarks, cross-bedding, etc. The Hutuo Group is characterised by thick carbonate and silicified rocks and has been divided into three subgroups. At the base is the Doucun Subgroup, which is dominated by terrigenous clastic sediments. This is overlain by the Dongye Subgroup, which is characterised by claret sandstone or slate in the lower part, and transitions upward into interbedded sandstone and carbonate with Stromatolites. It is dominated by dolomitic carbonate in its upper part. This is unconformably overlain by the Goujiazhai Subgroup, which consists of sandstone and local conglomerates.
The Hutuo Group was previously determined to be more than 2366 million years old, based zircon Uranium-Lead ages (when zircon forms it often contains trace amounts of uranium, which decays into, amongst other things, lead at a
known rate; since lead will not have been present in the original crystal matrix, it is possible to calculate the age of a zircon from the
ratio between these elements) from metamorphosed basic volcanic lava. More recent zircon age estimates made with more sensitie techniques are significanty younger, typically between 2150 and 1950 million years old.
Yin et al. investigated the Wenshan, Hebiancun, and Tianpengnao Formations of the Dongye Subgroup of the Hutuo Group for microfossils. The Wenshan Section is located 17.6 km southwest of Wutai County (38°36′ North and 113°7′ East). It comprises the lowermost Wenshan Formation, which is about 500 m thick and composed of metamorphic slate, and in consideration lithology of the single; 7 rock samples of slate were collected from the upper part of this section. The overlying Hebiancun Formation, which is about 1400 m thick and contains well-developed silicified carbonates containing Stromatolites and chert-like concretions. In order to obtain more possible preservation of fossil material from this deposited sequence, especially those of silicified Stromatolites and chert-like concretions, 59 rock samples were collected from middle-upper part of the Hebiancun Formation. In addition, and 76 phyllite rock samples from the uppermost Tianpengnao Formation of the Dongye Subgroup were collected from the Geziling Section, which is about 4 km east of Wutai County (38°44′ North, 113°17′ East).

Stratigraphic column of the Palaeoproterozoic Hutuo Group and sampling horizons of the Dongye Subgroup. Yin et al. (2020).
Standard palynological maceration was mainly used to obtain organic-walled microfossils from 12 slate samples of the Wenshan Formation and 76 phyllite samples from the Tianpengnao Formation. Samples of 50 g were cleaned and macerated with hydrocholric acid (37%) and hydrofluric acid (40%). Organic residues were either concentrated by heavy liquid with a specific gravity of 2.1–2.2, or poured through a 10 μm nylon mesh, and fixed slices were prepared with Canada balsam for the mounting medium of slices and sealed by paraffin. To obtain more fossil material and checking out possible contamination, which was mainly aimed at the phyllite of the Tianpengnao Formation, repeated macerations were performed. In result, total 14 samples including 2 slate samples of the Wenshan Formation and 12 phyllite samples of the Tianpengnao Formations produced organic-walled microfossils.
Additionally, 5 rock thin sections of phyllite of the Tianpengnao Formation (at least 6 mm in thickness) were cleared in distilled water were etched in dilute 8% hydrofluoric acid for 2–3 min, then cleared with distilled water, which were repeated processing in six times. Such etched rock thin sections were observed under scanning electron microscope to show preserved specimens in situ.
Outcrop photographs of the Tianpengnao Formation in the Geziling section and the Hebiancun Formation in the Wenshan section. (A), (B) Black phyllite or greyish-green phyllite interbedded with carbonates in the lower (A) and upper (B) parts of the Tianpengnao Formation. (C) Siliceous concretions in dolostone of the Hebiancun Formation. Yin et al. (2020).
The Tianpengnao Formation (at least 1.95 billion years old), which is the the uppermost formation of the Dongye Subgroup contains a higher diversity of organic-walled microfossils than the underlying formations. Organic-walled microfossils from the Tianpengnao Formation were mostly obtained through palynological preparation of phyllite. They were strongly carbonized and appeared as opaque vesicles. Individual specimen showed wall folds with a few fine spines under the scanning electron microscope. Similar opaque specimens in situ also found in thin sections. of fuchsia phyllite from the lower part of the Tianpengnao Formation. However, greyish-green phyllite from the upper part of the Tianpengnao Formation yielded much better preserved organic-walled microfossils.
The microfossils found by Yin et al are all considered to be Acritarchs or Cyanobacteria.
Acritarchs
are unicellular Eukaryotic organisms (organisms with cells with a
discrete nucleus) that appear in the fossil record from about 3200
million years ago until the end of the Permian, and possibly later (depending on what is classified as an Acritarch).
They're affinities are unclear, and the group is probably paraphyletic
(not all members sharing a common ancestry), though the majority are
thought to have been unicellular planktonic Algae or the resting cysts
of other unicellular organisms.
Cyanobacteria are filament-forming photosynthetic Bacteria found across
the globe and with a fossil record dating back over 3.5 billion years.
They are thought to have been the first organisms on Earth to obtain
carbon through photosynthesis, and it is also thought that the
chloroplasts (photosynthetic organelles) of eukaryotic plants and algae
are descended from Cyanobacteria that lived symbiotically within the
cells of ancient eukaryotes. Cyanobacteria are often known as Blue-Green
Algae, but this is somewhat misleading, as the term Algae is otherwise
restricted to photosynthetic eukaryotes (no other group of
photosynthetic Bacteria are referred to as Algae), and because not all
Cyanobacteria are blue-green in colour; many are dark green or even
black.
The first Acritarch described by Yin et al. is assigned to the genus Dictyosphaera, but not to species level. A single specimen was obtained from a siliceous lens in a dolostone containing Stromatolite from the Hebiancun Formation, at the Wenshan Section. This is a spheroidal vesicle, thin-walled, with very fine net-like ornamentation on its surface, forming a polygonal or subrounded mesh with 3–6 μm in diameter; the vesicle diameter is about 52 μm; no excystment structure was observed.
The Achritarch Dictyosphaera sp., in a in thin sections of siliceous material obtained from the Wenshan Section of the Hebiancun Formation. Scale bar is 10 μm. Yin et al. (2020).
The second Acritarch described is placed on a new species and genus, and given the name Dongyesphaera tenuispina, where 'Dongyesphaera' means 'sphere from Dongye' and 'tenuispina' means 'fine-spined'. Six specimens of this Acritarch were obtained by palynological maceration of material from the upper part of the Tianpengnao Formation. They are spheroidal to sub-spheroidal vesicles, the walls of which are psilate (lacking in ornamentation) and prominently wrinkled; with fairly short, conical processes or protrusions of varying length (0.5–4.2 μm, typically 0.8–1.5 μm), their termination showing as round; vesicles are 30–35 μm in diameter; no excystment opening was observed.
Dongyesphaera tenuispina, obtained from the upper greyish-green phyllite of the Tianpengnao Formation in the Geziling Section by palynological maceration. Scale Bars are 10 μm. Yin et al. (2020).
Acritarchs of the genus Leiosphaeridia were found in both palynological maceration and thin section from phyllite of the Tianpengnao Formation, siliceous lenses in dolostone containing Stromatolites of the Hebiancun Formation, and slate of the Wenshan Formation. spp. were obtained by palynological maceration and thin section. Most are strongly carbonized and even opaque. Some specimens obtained from phyllite of the upper part of the Tianpengnao Formation are less carbonized. None of these are assigned to species level by Yin et al. The specimens are spheroid vesicles, with a circular outline in compressed specimens; the wall surface typically is psilate or with inconspicuous ornament; some specimens show irregular folds; the vesicle diameter is 33–65 μm; no excystment structure was observed.

Leiosphaeridia spp., (A) obtained by palynological maceration from the upper greyish-green phyllite; (C) and (F) obtained by palynological maceration from the lower amaranth phyllite; (J) found in a thin section of the lower amaranth phyllite. Yin et al. (2020).
A single specimen is assigned to the genus Satka. This is a spherical colony-like specimen aggregated with many cell-like spheroids that has been compressed and appears to be enveloped by a thin outer membrane. The included spheroids are deformed and show different shapes and sizes. Single spheroids are 3–7 μm in diameter, and the whole vesicle about 40 μm in diameter. This specimen comes from a greyish-green phyllite of upper part of the Tianpengnao Formation, from the Geziling location.
Satka sp., from the upper greyish-green phyllite. Yin et al. (2020).
The first Cyanobacteria described by Yin et al. are assigned to the species Eoentophysalis belcherensis. These are irregular clusters that contain small and large spheroidal to sub-spheroidal cell-like units singly or in pairs; they are in a crowded arrangement, with a common thin envelope. Cell-like units are typically 1.8–2.5 μm in diameter; irregular clusters are 15–25 μm across and are frequently stuck together. These were foiund in thin sections of siliceous lenses in dolostone containing Stromatolites from the Hebiancun Formation.
Eoentophysalis belcherensis, from the Hebiancun Formation. Scale bar is 10 μm. Yin et al. (2020),
Seven specimens found in thin sections of two samples from siliceous concretions in dolostone of the Hebiancun Formation are assigend to a new species of Eoentophysalis; which is given the specific name hutuoensis, meaning 'from Hutuo', in reference to the Hutuo River in the Wutaishan area of Shanxi Province, China. Eoentophysalis hutuoensis comprises cell-like units spheroidal, ellipsoidal or deformed by compression, and 2.5–12 μm in diameter; mostly single, a few in possible pairs and irregular clusters; characteristically crowded in colonies which are enveloped by a thick sheath-like material. Therse are several hundred cell-like units arranged in clusters or extensive colonies, that are enveloped by an opaque sheath-like material (about 5–8 μm thick) and characterized by suborbicular holes (at least 1 μm in diameter); nearly all cell-like units are single, and more distinct in laser scanning confocal microscope images. Some of show ‘lining structure’ (or remains of plasmolysis). Clusters or colonies 78–126 μm across.

Eoentophysalis hutuoensis under laser scanning confocal microscope. Yin et al. (2020).
The third Cyanobacterium described by Yin et al. is assigned to the species Sphaerophycus medium. A single specimen from a siliceous lens in a dolostone containing Stromatolite of the Hebiancun Formation comprises an irregular clump of cells about 135 μm long and 86.5 μm wide; the cells generally not mutually deformed. Individual cells are spheroidal and ellipsoidal and 4.5–12.5 μm in diameter; cell walls are about 0.5 μm thick.
Sphaerophycus medium, from a siliceous lens in a dolostone containing Stromatolite of the Hebiancun Formation. Yin et al. (2020).
The fourth Cyanobacterium described is placed in the genus Pseudodendron, but not assigned to species level. This comprises a single compressed organic carbon specimen that has two short branches connected with a single, main ‘tube-like’ filament along two sides; the surface is nearly psilate and no preserved cellularity or outer enveloped material is observed. The ends of both branches and the main filament are truncated. The main filament 140 μm long, and 5–8 μm wide; the branches 20–48 μm long and 4–4.5 μm wide. Since only one incomplete specimen was found, its placement in Pseudodendron is tentative. The specimen was observed in a thin sections of siliceous lens from a dolostone containing Stromatolite of the Hebiancun Formation from the Wenshan Section.
Pseudodendron sp., from a thin sections of siliceous lens from a dolostone containing Stromatolite of the Hebiancun Formation from the Wenshan Section. Yin et al. (2020).
The final Cyanobacteria described is Siphonophycus kestron. Many specimens of this were found in thin sections of dolostone containing Stromatolites of the Hebiancun Formation. They are single filamentous microfossils, unbranched, nonseptate, surface smooth; only preserved as carbon membrane-like remains due to degradation; and 5–8 μm across.
Specimen of Siphonophycus kestron, from a thin section of dolostone containing Stromatolite of the Hebiancun Formation. Yin et al. (2020).
The Paleoproterozoic (2.5-1.6 billion years ago) was a critical period in Earth’s evolution. During it, important global events, such as glaciation, atmospheric oxygenation over the early period, and the following Lomagundi-Jatuli isotopic event occurred.
The Hutuo Group, as a typical Palaeoproterozoic sequence in the North China Craton, is distributed through the Wutai and Luliang mountains in Shanxi Province, China. The lithological character of the group is characterized by metamorphic deposits and volcanic rocks. Based on recent uranum-lead isotopic age dating, the Hutuo Group is constrained to the period between 2.14 and 1.95 billion years ago. Carbon isotope excursions in the Hutuo Group have been documented as a response to the Palaeoproterozoic global glaciation. Glaciogenic diamictite (a type of lithified sedimentary rock that consists of nonsorted to poorly sorted terrigenous sediment) has also been discovered in the Shijiazhuang Formation of the Hutuo Group at Wutai Mountain, North China which suggests a locally protracted glacial event could have extended to the Wutaishan area. Based on the uranium-lead isotopic ages and chronological framework of the Hutuo Group, a three-stage evolution in the Carbon¹³ isotope curve has been recognized in carbonates of the Hutuo Formation. The lower to middle part of the Dongye Subgroup shows oscillating positive and negative Carbon¹³ values that range between −5.2 and +2.7 parts per thousand relative to the Pee Dee Belemnite Standard (an increase in the proportion of Carbon¹³ relative to Carbon¹² is often indicative of an increase in photosynthesis, as photosynthetic organisms preferentially extract Carbon¹² from the atmosphere, whereas carbonate forming ones incorporate both in in a proportion reflecting atmosphere composition).
The microfossils from the Wenshan and Hebiancun Formations would be the fossil records of the geological period manifested by the aftermath of a positive excursion of Carbon¹³ (the Lomagundi-Jatuli isotopic event). The transition from an abnormally high organic carbon burial rate to massive oxidation of organic matter. The microfossils from the uppermost part of the Dongye Subgroup, (i.e. the Tianpengnao Formation), would represent the remains of Microphytoplankton during the geological period characterized by fluctuating Carbon¹³ levels.
The Palaeoproterozoic Hutuo Group was deposited in supra-tidal to sub-tidal environments. Furthermore, the Dongye Subgroup was followed by a remarkable transition of geochemical environments. In such palaeoenvironments, phosphates deposited on the continental margin of North China during late early Palaeoproterozoic. In the middle of the Hebiancun Formation, phosphatic deposits developed in association with dolomitic carbonates and a few terrigenous clasts in the studied Wenshan section. Several phosphatized specimens of spheroidal and filamentous Cyanobacteria and Leiosphaeridia-like forms were found in the phosphatic horizon. Typically, these specimens are poorly preserved, possibly due to late oxidation. Some specimens of Eoentophysalis hutuoensis were preserved as compressed carbon membranes. Such well-preserved Palaeoproterozoic microfossils, especially Eoentophysalis as multicellular colonies, have rarely been reported before. To understand their detailed morphological structure and elemental composition, scanning electron microscope associated with energy spectrum test was used for individual specimen of Eoentophysalis hutuoensis. Many sub-spherical individual cells were embedded within or enveloped by a Carbon membrane. The main elements detected were Carbon, Silicon, Calcium and Magnesium. This could suggest that Cyanobacteria colonies were primitively buried in the dolomitic carbonate and silicification during diagenesis resulted in their preservation. Common early diagenetic silicification was observed in the carbonates as distinct chert layers or concretions intercalated within dolostones of the Hebiancun Formation.

Eoentophysalis belcherensis, from the Hebiancun Formation. Scale bar is 10 μm. Yin et al. (2020),
A few degraded coccoid and filamentous cyanobacteria microfossils have previously been reported from the Hebiancun Formation. Some poorly preserved organic-walled microfossils obtained by palynological maceration have been described from the Doucun Subgroup. Some of those specimens that showed triangular, polygonal and boat-shaped forms that were plausibly interpreted as being like Eukaryotic Protists. In morphological feature discrimination, those specimens probably resulted from taphonomic alteration or were contaminants. Eukaryotic microfossils, except multicellular forms, are normally characterized by a Eukaryotic cytoskeleton and endomembrane system, morphogenetic characters like a multilayered wall, distinct surface ornamentation, and excystment by partial rupture or a circular opening. The oldest fossil evidence for Eukaryotic Protists (e.g. Tappania and other ornamented forms) have been documented from about 1.41 billion years ago elsewhere in the world. The new genus Dongyesphaera described by Yin et al. from the Tianpengnao Formation has a distinct fine spinous ornament on the vesicle wall, which would be recognized as eukaryotic protist. Additionally, a specimen identified as Dictyosphaera sp. was found in the Hebiancun Formation. The morphological genus Dictyosphaera has mostly been described from Late Palaeoproterozoic to Mesoproterozoic sediments in China, Australia and America. It is characterised by a multilayered vesicle wall, polygonal network ornamentation and possible excystment structure and is interpreted as a Eukaryotic Protest. At present, just one specimen identified as Dictyosphaera sp., by displaying network ornamentation on its vesicle surface, was found in the Hebiancun Formation, with an age of approximately 2010 million years. This suggests that the Eukaryotic Protist exercised metabolic activities rarely observed to have occurred in the early Palaeoproterozoic ocean. Up to the middle Palaeoproterozoic sequence, more specimens ornamented with fine conical spines, named as Dongyesphaera tenuispina, occurred in the greyish-green phyllite of the upper part of the Tianpengnao Formation (aged at about 1950 million years). Additionally, in siliceous lenses of dolostone containing stromatolite in the Hebiancun Formation, many Coccoid Cyanobacteria, such as Eoentophysalis and Eogloeocapsa were preserved, and some squashed Leiosphaerids, typically over 50 μm in diameter, have been preserved in situ. No obvious surface ornament was observed. However, the occurrence of the individual specimen of Dictyosphaera in the Hebiancun Formation implies that possible Eukaryotic organisms already existed around 2026 million years ago. Therefore, the microfossil evidence from Yin et al.'s study suggests that Eukaryotic organisms occurred earlier than, at least 2000 million years ago and quite lower morphological diversity of Eukaryotic organisms at the geological epoch. The recent discovery of early Precambrian microfossils, e.g. fungus-like mycelial fossils from 2.4 billion-year-old basalts of the Ongeluk Formation in South Africa, could suggest that Eukaryotic organisms may have occurred earlier than previously thought.
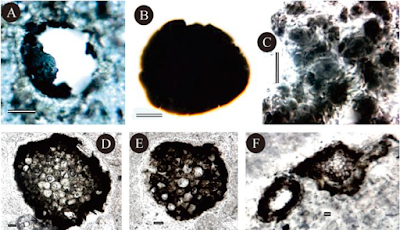
Photomicrographs of microfossils from the Hebiancun and Wenshan Formations in the Wenshan section. (A), (B) Leiosphaeridia sp., (A) broken specimen in thin section of silicified slate; (B) obtained by palynological maceration. (C) Degraded Coccoidal Cyanobacterium-like aggregation in a thin section of flint within crystalline dolostone, (D), (E) Eoentophysalis hutuoensis, in a thin section of a siliceous concretion in dolostone. Yin et al. (2020).
On the basis of microfossils found in samples from the Dongye Subgroup of the Palaeoproterozoic Hutuo Group in the Wutai Mountains of Shanxi Province, China, Yin et al. conclude the following: (1) Based on published geological data, there was an increased influence of oxygen on the carbon cycle during deposition of the Dongye Subgroup. For instance, phosphatised microfossils in phosphatic deposits of the Hebiancun Formation underwent stronger oxidation and show indistinct morphological aspects. (2) Owing to a remarkable increase in oxygen during the early Palaeoproterozoic, Eukaryotic Protists exercised metabolic activities rarely occurred in suitable environment, although Cyanobacteria were dominant in the early Palaeoproterozoic ocean. (3) The morphological diversity of Cyanobacteria appears to increase after about 2.0 billion years ago, especially the occurrence of many colonylike forms, such as the morphological genera Eoentophysalis and Sphaerophycus. Follow on research of microfossil records obtained from terrestrial clastic sediments of lower part of the Hutuo Group and greenschist of Neoarchean ‘Gaofan Subgroup’ is a potential project to reveal evolution of Earth’s life before and after global glaciation event.
See also...