Determining when groups of organisms first originated is one of the great challenges faced by evolutionary biologists. Traditionally, the fossil record has been the only way to determine when such groups first appeared, although since the 1960s molecular clock techniques, which use mutation rates to determine when groups of organisms diverged, have also been employed. Modern studies typically use a combination of the two techniques to achieve better results, but the accuracy of any study is still dependent on the quality of the available data.
The fossil record is not complete. Organisms stand a far better chance of being preserved in some environments that others, and organisms themselves differ greatly in their potential to leave fossil remains. Furthermore, Humans are often selective in both where they look for fossils, and in the types of fossils they collect, concentrating on larger, more eye-catching fossils, and often overlooking unconventional deposits which do not produce abundant, obvious fossils completely. Fossil groups which have skeletons made up of large numbers of small sections, such as small Vertebrates and Echinoderms, are often overlooked in environments which do not produce articulated skeletons, leaving gaps in our knowledge of their histories, even though their fossils are both numerous and widespread. Deposits from environments where articulated skeletons are unlikely to be preserved, such as the deep marine seafloor (where sedimentation rates are very low, and storm events never rapidly cover organisms) are often overlooked in palaeontological studies. These environments, however, often contain numerous disarticulated skeletal elements, which can potentially tell us a great deal about the history of traditionally undersampled groups.
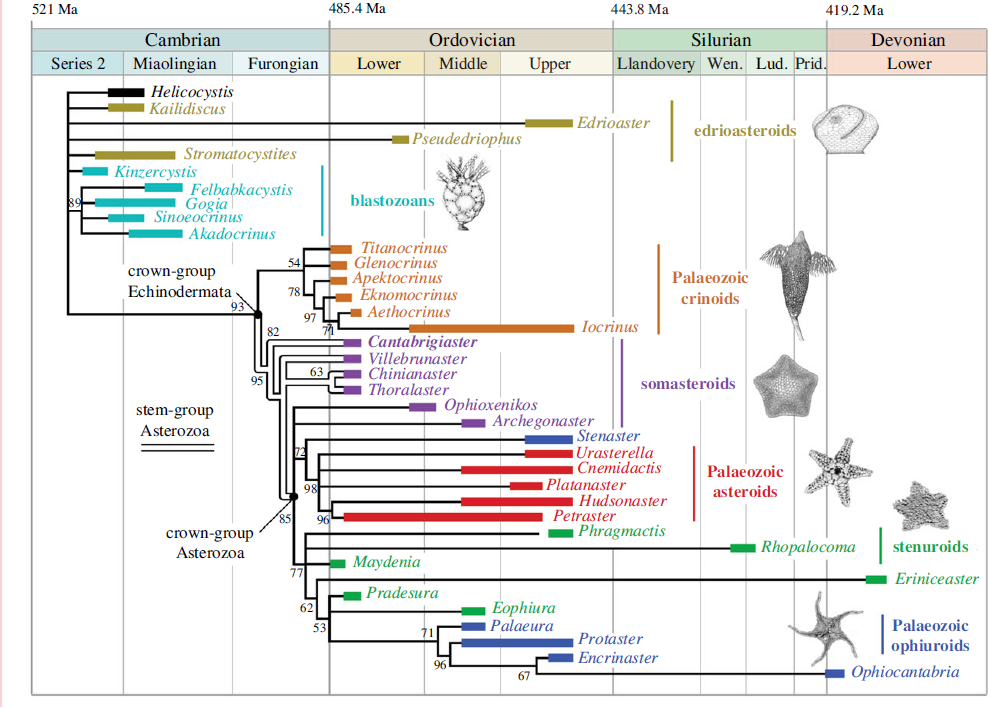
Ophiuroids, or Brittle Stars, are the most specious of the five extant classes of Echinoderms, and a well resolved phylogeny, based upon molecular data. However, the skeleton of Ophiuroids typically disaggregates into a large number of sand-sized particles within a few hours of death, with the result that fossils of entire Ophiuroids are very rare. The plates which make up the skeletons of Ophiuroids are actually very distinctive, making it possible to identify individual plates to the species level, and a great deal of work has been done on Mesozoic and Cainozoic Ophiuroids using individual plates, but the Palaeozoic Ophiuroid fossil record has been largely overlooked to date.
In a paper published in the journal Geology on 3 January 2023, Ben Thuy of the Department of Palaeontology at the Natural History Museum Luxembourg, Larry Knox of Earth Sciences at Tennessee Tech University, Lea Numberger-Thuy, also of the Department of Palaeontology at the Natural History Museum Luxembourg, and Nicholas Smith and Colin Sumrall of the Department of Earth and Planetary Sciences at the University of Tennessee, present the results of a study of Ophiuroid elements from deep water sediments from the Carboniferous of Oklahoma.
Thuy et al. examined 81 sieved micropalaeontological assemblages, obtained from bulk samples collected at Dutton Ranch in central southern Oklahoma for Ophiuroid samples. One of these, collected in 1981 from a blue-to-olive-grey shale identified as Unit 9BC, which is considered to be roughly equivalent to the Bostwick Member of the Lake Murray Formation, which dates to the latest Bashkirian (about 315 million years ago) proved to be particularly rich in Ophiuroid fossils. This unit was deposited in the northern Ardmore Basin, a northwestern extension of the Panthalassic Ocean. Based upon grain size, mineralogy, and Ostracod fauna, this deposit is thought to have been laid down in a deep slope or upper continental shelf environment.

In total, the samples produced about 400 Ophiuroid microfossils, including various types of arm plates (single ambulacrals and pairs fused into vertebrae; and lateral, ventral, and dorsal arm plates) as well as disc plates (radial shields and oral plates). From these lateral arm plates were chosen for further examination, as these are considered to show the greatest complexity, and are therefore the most useful taxonomically. Five distinct types of lateral arm plate were present in the sample, which Thuy et al. believe represent five separate species. Furthermore, two of these five species appear to be crown group Ophiuroids (the 'crown-group' of a group of organisms includes all living species within that group, their most recent common ancestor, and everything descended from that ancestor), assignable to extant orders, which Thuy et al. refer to as types A and B.

Type A lateral arm plates are elongate with coarsely reticulate stereom on the outer surface and a vertical row of large, freestanding spine articulations on an elevated distal edge. The spine articulations are composed of a single opening encompassed by a pair of arched dorsal and ventral lobes forming a lens-shaped elevation. The inner side has a low and poorly defined vertebral articular knob and a large tentacle notch. The general shape of these plates, as well as the shape of their arm articulations and the presence of coarse reticulation on their outer surfaces is consistent with the modern Ophiuroid order Ophioscolecida, and possibly with the genus Ophioscolex, although Thuy et al. do not go as far as assigning the specimens to genus level.
Type B lateral arm plates are by far the most common in the assemblage, making up about 90% of all the lateral arm plates collected. These are robust and strongly arched, and it appears likely that a pair of them would have been capable of completely enclosing the vertebrae of the arm. The outer surface ornamentation comprises a fine tuberculation, proximally bordered by a band of more coarsely meshed stereom including a central area with a fine horizontal striation and two spurs that establish the position of articulation with the overlapping adjacent lateral arm plate. The inner side of the lateral arm plate has a single, vertical ridge-shaped vertebral articular structure and a deep tentacle notch. The outer distal edge of the lateral arm plates bears a vertical row of small, freestanding spine articulations, each comprising a pair of parallel, horizontal dorsal and ventral lobes encompassing a small nerve opening and a slightly larger muscle opening. The type of spine articulation seen in these plates is exclusively seen in members of the Order Amphilepidida, which again is still extant today.
On the basis of their numeric domination of the sample, other plates found in the Dutton Ranch assemblage can be assumed to have come from the same species as the Type B lateral arm plates. These include ventral arm plates (which show similar ornamentation to the lateral arm plates) and radial shields, although Thuy et al. caution that, although the ventral arm plates appear to be consistent with the lateral arm plates, they would not, in themselves, be sufficient to assign a fossil to the Order Amphilepidida.
The placement of at least two species of Ophiuroids from the Dutton Ranch assemblage within modern orders is surprising, as this implies that these modern orders had emerged considerably before the theoretical date given for the emergence of the crown group Ophiuroids by molecular clock analysis. To test this hypothesis, Thuy et al. performed a Bayesian phylogenetic analysis using a previously established list of significant characters for the group, and using the early Carboniferous stem-group Ophiuroid Aganaster gregarius as an outgroup. This phylogeny confirmed the Type A lateral arm plates as having come from a member of the Order Ophioscolecida, which was close to the modern genera Ophioscolex and Ophiolycus, while the Type B lateral arm plates as having come from a basal member of the Order Amphilepidida.

This phylogenetic analysis confirms that lateral arm plate types A and B represent the earliest known examples of the orders Ophioscolecida and Amphilepidida, as well as (collectively) the oldest known examples of the Superorder Ophintegrida. This in turn implies that not only had the crown group Ophiuroida arisen by 315 million years ago, it had had time to split into the two superorders, Ophintegrida and Euryophiurida, and that the Superorder Ophintegrida had time to split into the orders Ophioscolecida and Amphilepidida. It has previously generally been assumed that the modern Ophiuroid orders arose during the period of high biotic turnover following the End Permia extinction, which appears to be the case with other Echinoderm groups. Although a pre-End Permian radiation of modern Ophiuroids has been suggested previously, Thuy et al.'s study presents the first direct evidence of this.
As well as providing evidence for an earlier origin of the crown group Ophiuroids than has previously been expected, Thuy et al.'s study also challenges the generally held assumption that shallow shelf environments are the major driver of biological innovation in the oceans, with some groups subsequently spreading into deep-water environments, and instead adds to a growing body of evidence that deep marine environments can themselves be a source of innovation, producing groups of organisms that go on to invade the shallow seas.
The assumption that little biological innovation occurs in the deep marine environment may be linked to the paucity of fossil-producing deep-water sediments in the fossil record. This is partly because of the low sedimentation rates in these environments, which leaves the remains of any Animals which die there exposed on the surface for long periods, but is also linked to the recycling of the ocean seafloor, which makes unaltered pre-Mesozoic deep-water deposits extremely rare. Thuy et al.'s study also demonstrates that these obstacles can also be overcome, and that careful sampling of those deposits which have been preserved can uncover ghost-lineages of modern groups, adding to our understanding of evolution in deep time as well as deep marine environments.
See also...